





Transkript
BRAINFOOD
Die Bedeutung langkettiger, mehrfach ungesättigter Fettsäuren für die kognitive Entwicklung
JANINA WILLERS*, JAN PHILIPP SCHUCHARDT*, ANDREAS HAHN*
Janina Willers
Den langkettigen, mehrfach ungesättigten Fettsäuren (LC-PUFA, long chain polyunsaturated fatty acids) der Omega-3-(ω-3-) und der -6-(ω-6-)Reihe kommt in der prä- und postnatalen kognitiven Entwicklung bis ins Kleinkindalter eine zentrale Rolle zu. Besonders bedeutsam sind dabei die ω-6-Fettsäure Arachidonsäure sowie die ω-3-Fettsäure Docosahexaensäure. Sie beeinflussen als integrale Bestandteile von Membranphospholipiden die Struktur, Fluidität und Funktion neuronaler Membranen. Auf molekularer Ebene regulieren LC-PUFA die Transkription spezifischer Gene. Zudem fungieren sie als Präkursormoleküle für die Synthese von Eicosanoiden wie Prostaglandinen und Leukotrienen und nehmen dadurch Einfluss auf inflammatorische Prozesse in neuronalen Strukturen.
Struktur und Metabolismus von LC-PUFA
Mehrfach ungesättigte Fettsäuren der ω-3- und der ω-6-Reihe zählen bekanntermassen zu den für den Menschen zufuhressenziellen Nährstoffen. Stammsubstanzen der beiden Fettsäurefamilien sind die α-Linolensäure (ALA, 18:3 ω-3) beziehungsweise die Linolsäure (LA, 18:2 ω-6); der Mensch ist grundsätzlich nicht zur Synthese dieser Verbindungen befähigt. Ausgehend von ALA und LA werden endogen weitere hoch ungesättigte Fettsäuren wie Eicosapentaensäure (EPA, 20:5 ω-3), Docosahexaensäure (DHA, 22:6 ω-3) und Arachidonsäure (AA, 20:4 ω-6) im endoplasmatischen Retikulum vornehmlich der Leberzellen gebildet (Abbildung 1). Diesen LC-PUFA, die auch in präformierter Form mit der Nahrung aufgenommen werden, kommt ein Grossteil der biologischen Wirkungen der jeweiligen Fettsäureklasse zu. Die körpereigene Synthese von insbesondere EPA und DHA aus ALA
*Leibniz Universität Hannover, Institut für Lebensmittelwissenschaft und Humanernährung
erfolgt nur sehr langsam und in einem geringen Ausmass. Zusätzlich beeinflussen genetische, stressassoziierte sowie geschlechtsspezifische Faktoren die Effizienz der Konversion. So konnte beispielsweise gezeigt werden, dass die Umwandlung von ALA zu EPA und DHA bei jungen Frauen weitaus effizienter verläuft als bei jungen Männern (1). Auch die – der heutigen Ernährungsweise entsprechend – hohe Zufuhr von LA vermindert kompetitiv die Bildung von LC-PUFA (2). Wegen der üblicherweise hohen Aufnahme von AA mit der Nahrung ist die Versorgung mit dieser Fettsäure dennoch gesichert. Der Versorgungsstatus mit EPA und DHA hingegen gilt wegen der geringen Gehalte in der Nahrung und der skizzierten metabolischen Einschränkungen vielfach als zu gering.
LC-PUFA und fetale Entwicklung
Bereits während der ersten vier Schwangerschaftswochen entsteht im Prozess der Neurulation das Neuralrohr. Dabei differenzieren und vermehren sich die neuronalen Stammzellen in verschiedene
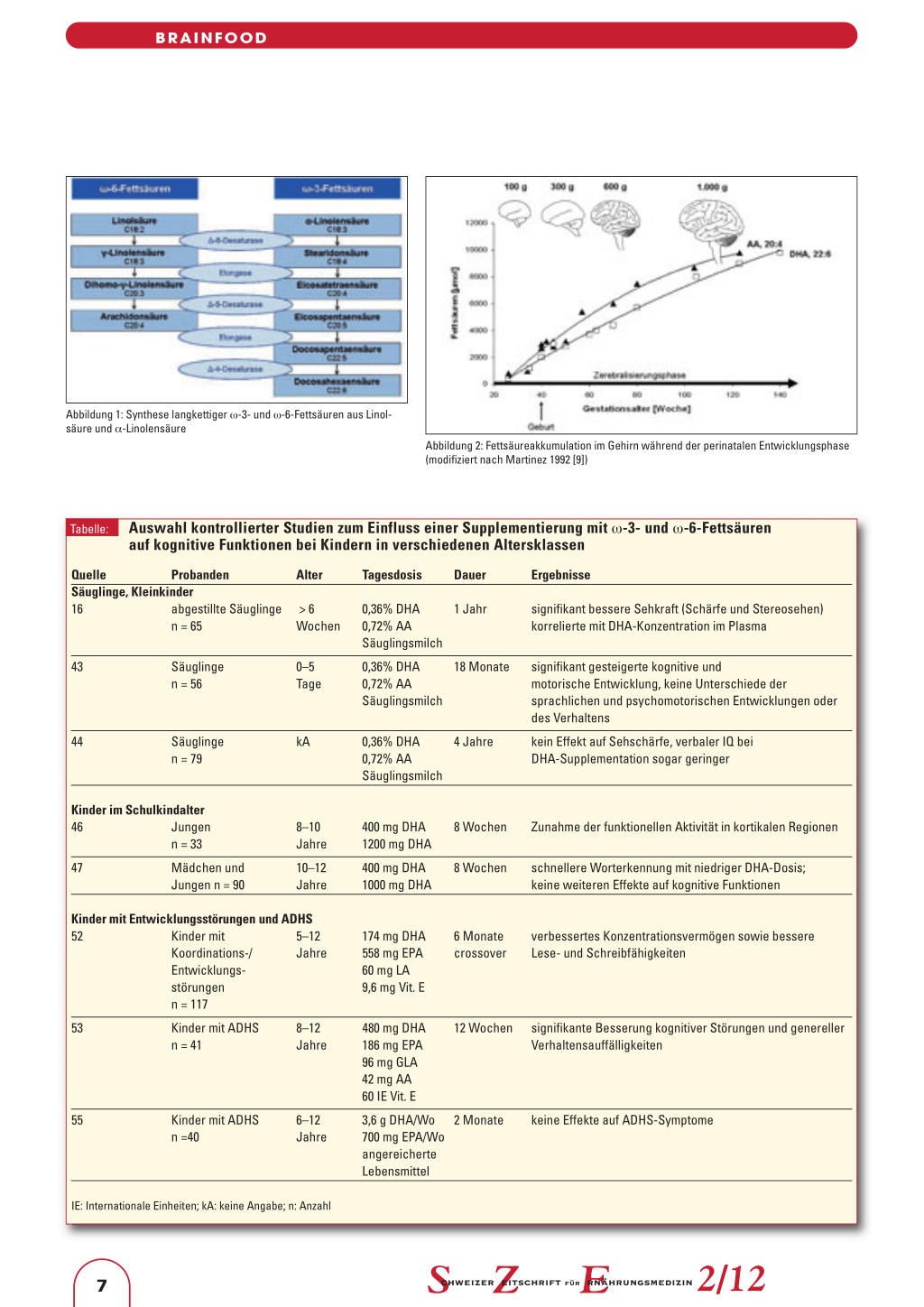
morphologische und funktionale Nervenzelltypen. Während des letzten Trimesters beginnt dann die umfangreiche Bildung von Synapsen (Synaptogenese) sowie die Myelinisierung (3, 4). In dieser Zerebralisierungsphase, die sich bis in die ersten Monate nach der Geburt erstreckt, wächst das Gehirn am schnellsten und reagiert am empfindlichsten auf einen Nährstoffmangel: Schwere nutritive Defizite in dieser Phase sind mit kognitiven und psychomotorischen Defiziten assoziiert (3, 5). Strukturell stellen AA und DHA die Schlüsselkomponenten neuronaler Membranen dar (Abbildung 2). Während AA in hohen Konzentrationen sowohl im Zentralnervensystem als auch in sämtlichen anderen Körpergeweben vorkommt, akkumuliert DHA verstärkt in der Netzhaut (Retina) und in der grauen Substanz der Grosshirnrinde (3). Insgesamt machen diese beiden Fettsäuren 15 bis 20 Prozent der Trockenmasse des Gehirns sowie mehr als 30 Prozent der Retina aus (6). Der Anteil anderer LC-PUFA im heranwachsenden Gehirn beziehungsweise in der
2/12
6
BRAINFOOD
Abbildung 1: Synthese langkettiger ω-3- und ω-6-Fettsäuren aus Linolsäure und α-Linolensäure
Abbildung 2: Fettsäureakkumulation im Gehirn während der perinatalen Entwicklungsphase (modifiziert nach Martinez 1992 [9])
Tabelle: Auswahl kontrollierter Studien zum Einfluss einer Supplementierung mit ω-3- und ω-6-Fettsäuren auf kognitive Funktionen bei Kindern in verschiedenen Altersklassen
Quelle
Probanden
Säuglinge, Kleinkinder
16 abgestillte Säuglinge
n = 65
Alter
>6 Wochen
43
Säuglinge
0–5
n = 56
Tage
44
Säuglinge
kA
n = 79
Tagesdosis Dauer
0,36% DHA
1 Jahr
0,72% AA
Säuglingsmilch
0,36% DHA
18 Monate
0,72% AA
Säuglingsmilch
0,36% DHA
4 Jahre
0,72% AA
Säuglingsmilch
Ergebnisse
signifikant bessere Sehkraft (Schärfe und Stereosehen) korrelierte mit DHA-Konzentration im Plasma
signifikant gesteigerte kognitive und motorische Entwicklung, keine Unterschiede der sprachlichen und psychomotorischen Entwicklungen oder des Verhaltens kein Effekt auf Sehschärfe, verbaler IQ bei DHA-Supplementation sogar geringer
Kinder im Schulkindalter 46 Jungen
n = 33
47 Mädchen und Jungen n = 90
8–10 Jahre
10–12 Jahre
400 mg DHA 1200 mg DHA
400 mg DHA 1000 mg DHA
8 Wochen 8 Wochen
Zunahme der funktionellen Aktivität in kortikalen Regionen
schnellere Worterkennung mit niedriger DHA-Dosis; keine weiteren Effekte auf kognitive Funktionen
Kinder mit Entwicklungsstörungen und ADHS
52
Kinder mit
5–12
Koordinations-/
Jahre
Entwicklungs-
störungen
n = 117
53
Kinder mit ADHS
8–12
n = 41
Jahre
55
Kinder mit ADHS
6–12
n =40
Jahre
174 mg DHA 558 mg EPA 60 mg LA 9,6 mg Vit. E
6 Monate verbessertes Konzentrationsvermögen sowie bessere crossover Lese- und Schreibfähigkeiten
480 mg DHA 186 mg EPA 96 mg GLA 42 mg AA 60 IE Vit. E
12 Wochen
3,6 g DHA/Wo 2 Monate 700 mg EPA/Wo angereicherte Lebensmittel
signifikante Besserung kognitiver Störungen und genereller Verhaltensauffälligkeiten
keine Effekte auf ADHS-Symptome
IE: Internationale Einheiten; kA: keine Angabe; n: Anzahl
7 2/12
BRAINFOOD
Netzhaut ist im Vergleich hierzu quantitativ gering. Innerhalb der Zellen finden sich die LC-PUFA vor allem als Bestandteil von Membranphospholipiden. Das Ausmass der Anreicherung der Fettsäuren im fetalen Gehirn via Plazentatransfer hängt in erster Linie von der Nahrungsaufnahme der Mutter ab und steigt mit der Zufuhr an. Obwohl die Plazenta und der fetale Organismus prinzipiell dazu in der Lage sind, die Fettsäurevorstufen (ALA und LA) in die längerkettigen Fettsäuren (EPA, DHA und AA) enzymatisch umzuwandeln, bestimmt vorwiegend die nutritive Zufuhr von DHA und AA die Akkumulationsrate im Gehirn (7, 8). AA ist entscheidend für das Wachstum und damit für die Massezunahme des Gehirns; bereits geringe Mängel sind mit einem erniedrigten Geburtsgewicht assoziiert (10). Demgegenüber sorgt DHA für eine normale kognitive und visuelle Entwicklung und konzentriert sich vor allem in hoch aktiven Membranen wie Synapsen und Fotorezeptoren (6). AA ist im mütterlichen Organismus normalerweise in mehr als ausreichender Menge vorhanden und wird dem Fetus entsprechend bereitgestellt. Die stark schwankende und oftmals marginale Versorgung mit DHA führt hingegen dazu, dass sich der Gehalt im maternalen sowie im fetalen Organismus und die geistige Leistung des Neugeborenen nutritiv beeinflussen lassen. Inzwischen liegen vielfältige Daten zur Bedeutung der DHA-Zufuhr für die kognitive Entwicklung vor.
Wirkungen von LC-PUFA auf die kognitive Entwicklung
Der Einfluss der LC-PUFA auf die kognitive Entwicklung und neuronale Funktionen ergibt sich durch ihre Beteiligung an zahlreichen physiologischen Prozessen (Abbildung 3).
Einfluss auf Struktur und Eigenschaften neuronaler Membranen Als integrale Bestandteile der neuronalen Membranphospholipide (12) beeinflussen LC-PUFA deren biophysikalische Eigenschaften und nehmen somit Einfluss auf ihre Funktionalität (13). Die Fluidität und damit beispielsweise auch die
Signalübertragung hän-
gen massgeblich von der
Zusammensetzung sowie
dem Anteil ungesättigter
Fettsäuren ab.
Eingebettet in die Phos-
pholipiddoppelschicht
von Membranen wirken
LC-PUFA zudem modulie-
rend auf lokale Proteine,
Rezeptoren und Ionenka-
näle. So zeigte sich in Tier-
versuchen, dass DHA die
Aktivität der Na+/K+-ATPase entscheidend beeinflusst (14).
Abbildung 3: Physiologische Funktionen von LC-PUFA in der neuronalen Entwicklung (modifiziert nach Schuchardt und Hahn 2011 [11]).
Insbesondere bei der Ent-
wicklung und Aufrechterhaltung des Seh- sowie Lern- und Gedächtnisleistung. So
vermögens spielen LC-PUFA als Mem- zeigten sich bei Kindern mit Aufmerk-
branbestandteile eine besondere Rolle samkeitsdefizit- und Hyperaktivitätssyn-
(15, 16). Membranständige Rezeptoren in drom (ADHS) Assoziationen zwischen Fett-
der Retina sorgen in Interaktion mit G- säureimbalancen und Störungen im
Proteinen und anderen Second-Messen- Neurotransmitterhaushalt (23). Ein Man-
ger-Systemen für die Transduktion neuro- gel der Neurotransmitter Serotonin und
naler Signale. Hier fördert insbesondere Dopamin führt bekanntlich zu Nervosität,
DHA die Funktion des G-Protein- Gedächtnisstörungen und motorischen
gekoppelten Systems in den Membranen Unruhesymptomen, die auch bei Kindern
der Fotorezeptorzellen und verbessert mit ADHS beobachtet werden (24).
auf diese Weise die Signalwege von
Metarhodopsin II (17).
Einfluss auf die Genregulation
LC-PUFA vermitteln ihre Funktionen im
Einfluss auf den Neurotransmitter-
Nervensystem auch durch Modulation der
haushalt
Expression spezifischer Gene (25–27). In
LC-PUFA, hier besonders ω-3-Fettsäuren, Rattenversuchen führte die Gabe von LC-
üben zudem einen Einfluss auf den Neu- PUFA (EPA und DHA) zu einer verstärkten
rotransmitterhaushalt aus. Bei Ratten Regulation von Genen, die mit synap-
konnte ein Zusammenhang zwischen tischer Plastizität, Zytoskelett und Mem-
dem ω-3-PUFA-Gehalt der Nahrung und branverband, Signalübertragung, Bildung
Veränderungen in der spezifischen Neu- von Ionenkanälen sowie dem Energie-
rotransmittersynthese gezeigt werden stoffwechsel assoziiert waren (28). Diese
(18). Ein chronischer Mangel an ω-3- Effekte scheinen unabhängig vom Einfluss
Fettsäuren verringerte dabei die Dopa- der Fettsäuren auf die Membranzusam-
minrezeptorbindung und erhöhte die mensetzung zu sein (29). Vermittelt wer-
Serotoninrezeptordichte im frontalen den sie durch Interaktion mit nukleären
Kortex von jungen und älteren Tieren Transkriptionsfaktoren. Bei einem dieser
(19–21). Umgekehrt führte eine erhöhte Faktoren handelt es sich um den Leber-X-
Nahrungszufuhr an ω-3-Fettsäuren bei Rezeptor (LXR α und β), der im Gehirn
Ratten zu einer 40-prozentigen Erhöhung exprimiert wird und eine Rolle bei der Li-
der Dopaminkonzentration im Frontal- pidhomöostase sowie bei neurodegenera-
kortex sowie zu einer Erhöhung der tiven Erkrankungen spielt (29, 30). Die
Dopaminrezeptorbindung (22).
Genexpression wird allerdings nicht nur
Der Einfluss von ω-3-LC-PUFA erklärt auch direkt durch LC-PUFA reguliert, sondern
deren Bedeutung für komplexe neuro- vermutlich auch durch sekundäre Lipid-
physiologische Vorgänge wie Verhalten mediatoren wie zum Beispiel Resolvine.
2/12
8
BRAINFOOD
Einfluss auf die Synthese von Eicosanoiden Aus LC-PUFA entstehen im Organismus oxidierte Fettsäuremetabolite (Oxylipine). Zur Oxylipinfamilie gehören unter anderem Eicosanoide (Prostaglandine [PG], Thromboxane [TX] und Leukotriene [LT]), die sich aus C20-Fettsäuren ableiten, sowie Resolvine, Protectine und Maresine (31, 32). Eicosanoide stellen die bislang am besten untersuchte Oxylipinfraktion dar. Im Allgemeinen sind die physiologischen Wirkungen der aus EPA gebildeten Eicosanoide (PGA3, TXA3, LTE5) günstiger als die der aus AA (PGE2, TXA2, LTA4) entstehenden Mediatoren. So wirken die entsprechenden Derivate aus ω-3-Fettsäuren auch vasodilatorisch und antiinflammatorisch. Neuere Daten deuten darauf hin, dass Resolvinen möglicherweise eine noch stärkere antiinflammatorische Wirkung zukommt als den jeweiligen Eicosanoiden (31). Die mechanistischen Grundlagen und Wirkweisen dieser und anderer Oxylipinderivate sind bisher noch nicht bekannt. Insbesondere DHA und ihre Oxidationsprodukte wirken über antineuroinflammatorische Regulationswege neuroprotektiv. Tierstudien konnten zeigen, dass ein induzierter Mangel an ω-3-Fettsäuren bei Ratten die DHA-Spiegel im Gehirn reduzierte, während die aus AA abgeleiteten Derivate anstiegen und damit proinflammatorische Signalkaskaden verstärkten (33, 34). Es wird daher vermutet, dass eine chronisch niedrige Aufnahme an ω-3-Fettsäuren die Auslösung entzündlicher Prozesse begünstigt. Möglicherweise können auf diesem Wege mit Neuroinflammation einhergehende Erkrankungen wie Morbus Alzheimer, Morbus Parkinson und Depressionen gefördert werden. Gleichzeitig ergeben sich hierdurch präventive und therapeutische Perspektiven für den Einsatz von ω-3Fettsäuren (33).
Untersuchungen zum Einfluss von LC-PUFA auf kognitive Funktionen Inzwischen liegen verschiedene Untersuchungen zum Einfluss von LC-PUFA auf kognitive Funktionen bei Säuglingen und
Kleinkindern sowie im Schulkindalter vor (Tabelle). Die dabei erzielten Ergebnisse aus Interventionsstudien sind uneinheitlich und teilweise widersprüchlich, was vermutlich auf methodische Mängel sowie erhebliche Unterschiede im Hinblick auf Studiendauer, Ausgangsstatus, Zusammensetzung und Dosierung der LC-PUFA zurückzuführen ist. Zudem ist umstritten, welche methodischen Ansätze im Einzelfall geeignet sind, kognitive Funktionen beziehungsweise Intelligenz differenziert zu erfassen (35). Beobachtungs- sowie Kohortenstudien kommen überwiegend zum Ergebnis, dass eine regelmässige Zufuhr von Fisch während der Schwangerschaft mit einer signifikanten Verbesserung der kognitiven kindlichen Entwicklung bis ins Kleinkindalter assoziiert ist (36, 37). Zudem erzielten Kinder in verschiedenen Interventionsstudien im Kleinkindalter eine im Vergleich zu Kontrollgruppen bessere visuomotorische Koordination sowie bessere Ergebnisse in kognitiven Scores, wenn ihre Mütter ab der 18. bis 20. Gestationswoche bis zur Entbindung ω-3-Fettsäuren in Form von Fischöl supplementiert hatten (38, 39). Allerdings finden sich auch Studien, die keinen Effekt einer pränatalen ω-3-Fettsäuresupplementierung auf kognitive Funktionen oder den globalen IQ bei Kleinkindern nachweisen konnten (40, 41). Ein 2012 veröffentlichtes systematisches Review (42) kommt zu dem Ergebnis, dass auf Basis der derzeit publizierten Studien keine anhaltende Verbesserung der kognitiven Entwicklung durch die pränatale Supplementierung mit ω-3-Fettsäuren bei Säuglingen belegt ist. Ein vergleichbar uneinheitliches Bild zeigt sich auch in Studien, die ω-3- und ω-6-Fettsäuren postnatal verabreichten. So führte eine postnatale Supplementierung von Neugeborenen sowie abgestillten Säuglingen mit DHA und AA über Säuglingsmilchnahrung in einigen Studien zu einer signifikant gesteigerten kognitiven und motorischen Entwicklung (16, 43). Andere Untersuchungen wiederum zeigten keine Veränderungen und sogar eine Verschlechterung im Hinblick auf den verbalen IQ (44). Unklar bleibt auch,
ob eine postnatale Supplementierung den gleichen positiven Effekt auf die kognitive Entwicklung haben kann wie eine pränatale Supplementierung während der Schwangerschaft (45). Vergleichsweise wenig Daten liegen bis anhin zum Einfluss von LC-PUFA auf die kognitiven Funktionen von Kindern im Schulalter vor. Bei gesunden Jungen im Alter von 8 bis 10 Jahren zeigte sich nach einer achtwöchigen ω-3-Fettsäuresupplementierung (400 mg/Tag oder 1200 mg/ Tag) mithilfe der Magnetresonanztomografie eine Zunahme der funktionellen Aktivität in kortikalen Regionen, die mit Aufmerksamkeit und Lernen in Zusammenhang stehen (46). Ergebnissen einer anderen Studie zufolge hatte eine achtwöchige DHA-Supplementierung (400 mg/ Tag oder 1000 mg/ Tag) bei 10- bis 12-jährigen gesunden Kindern jedoch keinen positiven Effekt auf die Gehirnfunktion. Lediglich ein kognitiver Parameter (Geschwindigkeit der Worterkennung) verbesserte sich in der DHA-Gruppe mit niedriger Dosierung. Dieses Ergebnis wurde von den Autoren aber als zufällige Schwankung angesehen und nicht als ein tatsächlicher kognitiver Effekt (47). Insgesamt kann, angesichts der zahlreichen Hinweise auf einen physiologischen Nutzen, eine gute maternale, prä- und postnatale sowie kindliche Versorgung mit DHA und AA als grundsätzlich sinnvoll angesehen werden. In welchem Masse und unter welchen Bedingungen die zusätzliche Gabe von insbesondere ω-3Fettsäuren mit einem zusätzlichen Nutzen verbunden ist, ist dabei differenziert zu betrachten und nicht abschliessend geklärt. Unstrittig ist, dass eine mangelnde Versorgung mit ω-3-Fettsäuren kognitive Entwicklungsdefizite begünstigen kann. Beispielsweise weisen Kinder, die in der Schwangerschaft nicht ausreichend mit ω-3-Fettsäuren versorgt waren, mehr Verhaltensprobleme sowie neuro-kognitive Störungen wie beispielsweise ADHS auf (6, 48, 49). Bei Kindern mit ADHS finden sich im Vergleich zu gesunden Kontrollen häufig erniedrigte Spiegel der Fettsäuren DHA, LA und AA in den Phospholipiden und Erythrozytenmembranen (50, 51).
9 2/12
BRAINFOOD
Inzwischen liegen Daten aus zahlreichen Interventionsstudien vor, in denen Kinder mit ADHS ω-3-Fettsäure-Supplemente erhielten (52–56). So verbesserten sich in verschiedenen Studien unter anderem das Konzentrationsvermögen sowie die Lese- und Schreibfähigkeiten nach Verabreichung von ω-3-Fettsäuren (EPA/DHA). Auch eine Metaanalyse konnte diese Resultate bestätigen. Die Ergebnisse von zehn plazebokontrollierten Studien mit insgesamt 699 Kindern deuten auf einen moderaten Effekt der ω-3-Fettsäuren, insbesondere EPA, bei den ADHS-Symptomen hin (54). Auch wenn die Effekte im Vergleich mit gängigen pharmakologischen ADHS-Therapien eher mässig ausfallen, spricht doch das geringe Nebenwirkungsprofil für eine Supplementierung mit ω-3-Fettsäuren. Insgesamt scheinen gesunde Kinder sowie Kinder mit ADHS im Kleinkind- und Schulalter von einer LC-PUFA-Supplementierung zu profitieren. Im Detail bleiben allerdings noch zahlreiche Fragen offen, so die nach den notwendigen Dosierungen sowie den erzielbaren Effektgrössen.
Zufuhrempfehlungen und Versorgungssituation
Die Empfehlungen verschiedener nationaler und internationaler Gremien für die Zufuhr von LC-PUFA für Erwachsene sowie für schwangere und stillende Frauen sind uneinheitlich. Da der Mensch prinzipiell dazu in der Lage ist, Vorläuferfettsäuren in die eigentlich bioaktiven hoch ungesättigten LC-PUFA umzuwandeln, beschränken sich die Zufuhrempfehlungen zumeist auf die beiden essenziellen Stammfettsäuren ALA und LA (57). Dieser Ansatz ist umstritten, da nachgewiesen werden konnte, dass die Umwandlungsrate, vor allem aus ALA in DHA, extrem niedrig und die interindividuelle Variabilität beträchtlich ist. Separate Empfehlungen für EPA und DHA erscheinen deshalb sinnvoll. Beispielsweise empfehlen die Referenzwerte für die Nährstoffzufuhr der Ernährungsgesellschaften in Deutschland, Österreich und der Schweiz (57) zur primären Prävention der koronaren Herzkrankheit eine Zufuhrmenge von 250 mg DHA und EPA pro Tag.
Zwischen der empfohlenen und der tatsächlichen Zufuhr herrscht häufig eine Diskrepanz, da der Anteil an ω-3-Fettsäuren in der vorherrschenden Nahrung moderner Industriestaaten verhältnismässig gering ist. Langkettige ω-3-Fettsäuren finden sich fast ausschliesslich in wenigen fettreichen Kaltwasserfischen wie Lachs, Makrele oder Hering. Aufgrund des geringen Gehalts an ω-3Fettsäuren in der üblichen Mischkost liegt die nutritive Aufnahme bei schwangeren Frauen häufig unter den Zufuhrempfehlungen (58, 59). Bei 20 untersuchten schwangeren kanadischen Frauen im zweiten und dritten Trimester erreichte die mittlere DHA-Zufuhr 82 ± 33 mg/Tag (59). Diese Situation ist vergleichbar mit anderen westlichen Ländern. Die mittlere tägliche Zufuhr an ω-3-Fettsäuren (EPA, DHA) von deutschen Frauen liegt gemäss Ernährungsbericht der Deutschen Gesellschaft für Ernährung (60) bei immerhin etwa 150 mg/Tag. Entsprechend der geringen Aufnahmemenge konnte darüber hinaus auch nur ein geringer Gehalt an ω-3-Fettsäuren in Frauenmilch festgestellt werden (61). Allerdings unterliegt beispielsweise der DHA-Gehalt in Frauenmilch einer grossen Schwankungsbreite. Untersuchungen ergaben, dass die DHAGehalte zwischen 0,06 und 1,40 Prozent der Gesamtfettsäuren liegen, wobei die höchsten DHA-Konzentrationen in Küstenregionen mit einem hohen Verzehr mariner Lebensmittel gefunden wurden (62). Die Versorgung mit ω-6-Fettsäuren bei schwangeren und stillenden Frauen in Industrieländern kann als gesichert angesehen werden, sodass keine Notwendigkeit einer erhöhten Aufnahme von AA während der Schwangerschaft gesehen wird, sofern eine bedarfsgerechte Zufuhr von LA vorliegt. In einem Konsenspapier verschiedener nationaler und internationaler Expertengruppen für perinatale Ernährung wurden auf Basis systematischer Literaturanalysen evidenzbasierte Empfehlungen zur Nahrungsfettzufuhr für schwangere und stillende Frauen ausgegeben. Demnach ist eine DHA-Aufnahme von mindestens 200 mg/Tag während der Schwangerschaft und in der Stillzeit anzustreben
(63, 64). Dies dient einer deutlichen Risikominderung für Frühgeburten und zur Förderung einer optimalen kognitiven und visuellen Entwicklung des Fötus und des Neugeborenen. Diese Zufuhrmenge kann durch den Verzehr von ein bis zwei Portionen fettreichem Seefisch pro Woche erreicht werden (65). Nach Angaben des Max-Rubner-Instituts ist bei dieser Zufuhrmenge die derzeitige Schadstoffbelastung (z.B. Quecksilber, Cadmium) der Seefische auf dem deutschen Markt als gering anzusehen. Für Kinder existieren derzeit noch keine Empfehlungen zur Zufuhr von EPA oder DHA. Nach Auffassung von Koletzko et al. (65) sind die bislang verfügbaren Informationen über den Einfluss von EPA und DHA bei Kindern zwischen 2 und 12 Jahren auf Wachstum, Entwicklung und Gesundheit nicht ausreichend, um daraus quantitative Zufuhrempfehlungen für Kinder ableiten zu können (66). Im Gegensatz zur menschlichen Muttermilch enthalten die meisten handelsüblichen Säuglingsmilchnahrungen keine nennenswerten Mengen an LC-PUFA, sofern keine Anreicherung erfolgt. Dies ist allerdings zunehmend der Fall, sodass einheitliche Zufuhrempfehlungen und Empfehlungen zur Anreicherung von Säuglingsmilch mit LC-PUFA sinnvoll und aus lebensmittelrechtlicher Sicht auch notwendig erscheinen.
Zusammenfassung und Schlussfolgerung
LC-PUFA sind von zentraler Bedeutung für Entwicklung und Funktion des Gehirns sowie des Nervensystems, vor allem während der kindlichen Entwicklung. Eine ausreichende Versorgung mit langkettigen ω-3- und ω-6-Fettsäuren (DHA, EPA, AA) ist daher für die normale Entwicklung während des fetalen Wachstums in der Zerebralisierungsphase sowie für die visuellen und kognitiven Funktionen bei Neugeborenen unentbehrlich. Bei einer üblichen westlichen Ernährungsweise ist die Zufuhr an ω-3-Fettsäuren – insbesondere EPA und DHA – stark limitiert. Sowohl die maternale als auch die fetale Versorgung ist unter Umständen von einem relativen ω-3-Fettsäure-Mangel betroffen.
2/12
10
BRAINFOOD
Die Beteiligung von LC-PUFA an physiologischen Prozessen innerhalb des Gehirns und des Nervensystems (Membranstabilität und -fluidität, Signalübertragung, Neurotransmitterhaushalt, Genregulation, Eicosanoidsynthese) erklärt, warum ein relativer Fettsäuremangel oder ein Ungleichgewicht zwischen ω-3- und ω-6Fettsäuren zu einer Beeinträchtigung der neuro-kognitiven Funktionen führen kann. Dies äussert sich bei Kindern beispielsweise in Form von Entwicklungsund Verhaltensstörungen. Inwieweit die kognitive Leistung von Kindern durch zusätzliche Gaben von LC-PUFA beeinflusst werden kann, ist derzeit noch nicht abschliessend geklärt. Verschiedene Expertengruppen empfehlen schwangeren und stillenden Frauen eine regelmässige Zufuhr langkettiger ω-3-Fettsäuren zur Förderung einer optimalen kognitiven und visuellen Entwicklung des Fetus und des Neugeborenen. Eine mittlere Aufnahme von mindestens 200 mg DHA pro Tag während der Schwangerschaft und Stillzeit sollte erreicht werden. Diese Zufuhrmenge kann im Prinzip durch den langzeitlichen Verzehr von ein bis zwei Portionen fettreichen Kaltwasserfisches pro Woche erreicht werden. In der Praxis dürfte es allerdings vielfach notwendig sein, auf entsprechende Supplemente zurückzugreifen. Inzwischen zeigt sich immer deutlicher, dass LC-PUFA, insbesondere der Omega3-Reihe, nicht nur für die Entwicklung der kognitiven Funktion in der frühen Lebensphase, sondern auch für deren Erhalt und die Prävention von Demenzerkrankungen im höheren Lebensalter von zentraler Bedeutung sind. Dieser Aspekt wird Gegenstand eines weiteren Beitrags sein.
Korrespondenzadresse: Dr. Janina Willers Leibniz Universität Hannover Institut für Lebensmittelwissenschaft und Humanernährung Am Kleinen Felde 30, D-30167 Hannover Tel. +49 (0)511-762 57 55 Fax +49 (0)511-762 5 729 E-Mail: willers@nutrition.uni-hannover.de
Interessenkonflikte: Es bestehen keine Interessenkonflikte. Die Autoren sind allerdings an Forschungskooperationen mit Unternehmen beteiligt, die unter anderem auch Produkte mit ω-3-Fettsäuren vertreiben.
Literatur: 1. Burdge GC, Calder PC. Conversion of alpha-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod Nutr Dev. 2005; 45 (5): 581–597. 2. Schuchardt JP, Hahn A. Essenzielle Fettsäuren und Hirnfunktion bei Kindern. Dtsch Apothek Z. 2008; 148 (10): 972–980. 3. Wainwright PE. Dietary essential fatty acids and brain function: a developmental perspective on mechanisms. ProcNutr Soc. 2002; 61 (1): 61–69. 4. Roth G, Strüber N. Pränatale Entwicklung und neurobiologische Grundlagen der psychischen Entwicklung. In: Cierpka (Hrsg.) Frühe Kindheit 0–3. SpringerVerlag, Berlin Heidelberg 2012; 4–20. 5. Uauy R, Dangour AD. Nutrition in brain development and aging: role of essential fatty acids. Nutr Rev. 2006; 64, 5: S24–S33. 6. Richardson AJ. Long-chain polyunsaturated fatty acids in childhood developmental and psychiatric disorders. Lipids. 2004; 39 (12): 1215–1222. 7. Marc I, Plourde M, Lucas M, Sterescu A, Piedboeuf B, Dufresne A, Nuyt AM, Lévy E, Dodin S. Early docosahexaenoic acid supplementation of mothers during lactation leads to high plasma concentrations in very preterm infants. J Nutr. 2011; 141 (2): 231–236. 8. Fidler N, Sauerwald T, Pohl A, Demmelmair H, Koletzko B. Docosahexaenoic acid transfer into human milk after dietary supplementation: a randomized clinical trial. J Lipid Res. 2000; 41 (9): 1376–1383. 9. Martinez M. Tissue levels of polyunsaturated fatty acids during early human development. J Pediatr. 1992; 120: S129–138. 10. Koletzko B, Braun M. Arachidonic acid and early human growth: is there a relation? Ann Nutr Metab. 1991; 35 (3): 128–131. 11. Schuchardt JP, Hahn A. Influence of long-chain polyunsaturated fatty acids (LC-PUFAs) on cognitive and visual development. In: Lifetime nutritional influences on cognition, behaviour and psychiatric illness. Woodhead Publishing Series in Food Sciences, Technology and Nutrition: Number 223. Woodhead Publishing Limited. 2011; 32–78. 12. Singh M. Essential Fatty Acids, DHA and Human Brain. Indian J Pediatr. 2005; 72 (3): 239–242. 13. Stillwell W, Wassall SR. Docosahexaenoic acid: membrane properties of a unique fatty acid. Chem Phys Lipids. 2003; 126 (1):1–27. 14. Turner N, Else PL, Hulbert AJ. Docosahexaenoic acid (DHA) content of membranes determines molecular activity of the sodium pump: implications for disease states and metabolism. Naturwissenschaften. 2003; 90 (11): 521–523. 15. Smithers LG, Gibson RA, McPhee A, Makrides M. Higher dose of docosahexaenoic acid in the neonatal period improves visual acuity of preterm infants: results of a randomized controlled trial. Am J Clin Nutr. 2008; 88: 1049–1056. 16. Birch EE, Hoffman DR, Castañeda YS, Fawcett SL, Birch DG, Uauy RD. A randomized controlled trial of long-chain polyunsaturated fatty acid supplementation of formula in term infants after weaning at 6 wk of age. Am J Clin Nutr. 2002; 75 (3): 570–580. 17. Litman BJ, Niu SL, Polozova A, Mitchell DC. The role of docosahexaenoic acid containing phospholipids in modulating G protein-coupled signaling pathways: visual transduction. J Mol Neurosci. 2001; 16 (2–3): 237–242.
18. Chalon S. Omega-3 fatty acids and monoamine neurotransmission. Prostaglandins Leukot Essent Fatty Acids. 2006; 75 (4–5): 259–269. 19. Zimmer L, Delion-Vancassel S, Durand G, Guilloteau D, Bodard S, Besnard JC, Chalon S. Modification of dopamine neurotransmission in the nucleus accumbens of rats deficient in n-3 polyunsaturated fatty acids. J Lipid Res. 2000; 41 (1): 32–40. 20. Zimmer I, Hembert S, Dward G, Breton P, Guilloteau D, Besnard J-C & Chalon S. Chronic n-3 polyunsaturated fatty acid diet-deficiency acts on dopamine metabolism in the rat frontal cortex: a microdialysis study. Neuroscience Letters. 1998; 240: 177–181. 21. Delion S, Chalon S, Guilloteau D, Besnard JC, Durand G. Alpha-Linolenic acid dietary deficiency alters age-related changes of dopaminergic and serotoninergic neurotransmission in the rat frontal cortex. J Neurochem. 1996; 66 (4): 1582–1591. 22. Chalon S, Delion-Vancassel S, Belzung C, Guilloteau D, Leguisquet AM, Besnard JC, Durand G. Dietary fish oil affects monoaminergic neurotransmission and behavior in rats. J Nutr. 1998; 128 (12): 2512–2519. 23. Frölich J, Döpfner M. Die Behandlung von Aufmerksamkeitsdefizit-/Hyperaktivitätsstörungen mit mehrfach ungesättigten Fettsäuren – eine wirksame Behandlungsoption? Z Kinder- und Jugendpsychiatrie und Psychotherapie. 2008; 36 (2): 109–116. 24. Gröber U. Hyperaktiv und hypermotorisch – ein Mangel an Methylphenidat? Mikronährstoffe bei ADHS. Zs. f. Orthomol. Med. 2008; 3: 5–8. 25. Berger A, Mutch DM, German JB, Roberts MA. Dietary effects of arachidonate-rich fungal oil and fish oil on murine hepatic and hippocampal gene expression. Lipids Health Dis. 2002; 1: 2. 26. Salvati S, Natali F, Attorri L, Di Benedetto R, Leonardi F, Di Biase A, Ferri F, Fortuna S, Lorenzini P, Sanchez M, Ricceri L, Vitelli L. Eicosapentaenoic acid stimulates the expression of myelin proteins in rat brain. J Neurosci Res. 2008; 86 (4): 776–784. 27. Jump DB, Clarke SD. Regulation of gene expression by dietary fat. Annu Rev Nutr 1999; 19: 63–90. 28. Kitajka K, Puskás LG, Zvara A, Hackler L Jr, Barceló-Coblijn G, Yeo YK, Farkas T. The role of n-3 polyunsaturated fatty acids in brain: modulation of rat brain gene expression by dietary n-3 fatty acids. Proc Natl Acad Sci USA. 2002; 99 (5): 2619–2624. 29. Kitajka K, Sinclair AJ, Weisinger RS, Weisinger HS, Mathai M, Jayasooriya AP, Halver JE, Puskás LG. Effects of dietary omega-3 polyunsaturated fatty acids on brain gene expression. Proc Natl Acad Sci USA. 2004; 101 (30): 10931–10936. 30. Wang L, Schuster GU, Hultenby K, Zhang Q, Andersson S, Gustafsson JA. Liver X receptors in the central nervous system: from lipid homeostasis to neuronal degeneration. Proc Natl Acad Sci USA. 2002; 99 (21): 13878–13883. 31. Zhang MJ, Spite M. Resolvins: Anti-Inflammatory and Proresolving Mediators Derived from Omega-3 Polyunsaturated Fatty Acids. Annu Rev Nutr. 2012; 32: 7.1–7.25. 32. Farooqui AA. n-3 fatty acid-derived lipid mediators in the brain: new weapons against oxidative stress and inflammation. Curr Med Chem. 2012; 19 (4): 532–543. 33. Orr SK, Bazinet RP. The emerging role of docosahexaenoic acid in neuroinflammation. Curr Opin Investig Drugs. 2008; 9 (7): 735–743. 34. Rao JS, Ertley RN, DeMar JC Jr, Rapoport SI, Bazinet RP, Lee HJ. Dietary n-3 PUFA deprivation alters expression of enzymes of the arachidonic and docosahexaenoic acid cascades in rat frontal cortex. Mol Psychiatry. 2007; 12 (2): 151–157. 35. Petermann F. Intelligenzdiagnostik. Kindheit und Entwicklung. 2006; 15 (2): 71–75.
11 2/12
BRAINFOOD
36. Mendez MA, Torrent M, Julvez J, Ribas-Fitó N, Kogevinas M, Sunyer J. Maternal fish and other seafood intakes during pregnancy and child neurodevelopment at age 4 years. Public Health Nutr. 2009; 12 (10): 1702–1710. 37. Oken E, Wright RO, Kleinman KP, Bellinger D, Amarasiriwardena CJ, Hu H, Rich-Edwards JW, Gillman MW. Maternal fish consumption, hair mercury, and infant cognition in a U.S. Cohort. Environ Health Perspect. 2005; 113 (10): 1376–1380. 38. Dunstan JA, Simmer K, Dixon G, Prescott SL. Cognitive assessment of children at age 2 (1/2) years after maternal fish oil supplementation in pregnancy: a randomised controlled trial. Arch Dis Child Fetal Neonatal Ed. 2008; 93 (1): F45–50. 39. Helland IB, Smith L, Saarem K, Saugstad OD, Drevon CA. Maternal supplementation with very-longchain n-3 fatty acids during pregnancy and lactation augments children’s IQ at 4 years of age. Pediatrics. 2003; 111 (1): e39–44. 40. Helland IB, Smith L, Blomén B, Saarem K, Saugstad OD, Drevon CA. Effect of supplementing pregnant and lactating mothers with n-3 very-long-chain fatty acids on children’s IQ and body mass index at 7 years of age. Pediatrics. 2008; 122 (2): e472–479. 41. Helland IB, Saugstad OD, Smith L, Saarem K, Solvoll K, Ganes T, Drevon CA. Similar effects on infants of n-3 and n-6 fatty acids supplementation to pregnant and lactating women. Pediatrics. 2001; 108 (5): E82. 42. Lo A, Sienna J, Mamak E, Djokanovic N, Westall C, Koren G. The effects of maternal supplementation of polyunsaturated fatty acids on visual, neurobehavioural, and developmental outcomes of the child: a systematic review of the randomized trials. Obstet Gynecol Int. 2012; 2012: 591531. 43. Birch EE, Garfield S, Hoffman DR, Uauy R, Birch DG. A randomized controlled trial of early dietary supply of long-chain polyunsaturated fatty acids and mental development in term infants. Dev Med Child Neurol. 2000; 42 (3): 174–181. 44. Birch EE, Garfield S, Castañeda Y, HughbanksWheaton D, Uauy R, Hoffman D. Visual acuity and cognitive outcomes at 4 years of age in a double-blind, randomized trial of long-chain polyunsaturated fatty acid-supplemented infant formula. Early Hum Dev. 2007; 83 (5): 279–284. 45. Hadders-Algra M. Prenatal and early postnatal supplementation with long-chain polyunsaturated fatty acids: neurodevelopmental considerations. Am J Clin Nutr. 2011; 94 (6 Suppl): 1874S–1879S.
46. McNamara RK, Able J, Jandacek R, Rider T, Tso P, Eliassen JC, Alfieri D, Weber W, Jarvis K, DelBello MP, Strakowski SM, Adler CM. Docosahexaenoic acid supplementation increases prefrontal cortex activation during sustained attention in healthy boys: a placebo-controlled, dose-ranging, functional magnetic resonance imaging study. Am J Clin Nutr. 2010; 91 (4): 1060–1067. 47. Kennedy DO, Jackson PA, Elliott JM, Scholey AB, Robertson BC, Greer J, Tiplady B, Buchanan T, Haskell CF. Cognitive and mood effects of 8 weeks’ supplementation with 400 mg or 1000 mg of the omega-3 essential fatty acid docosahexaenoic acid (DHA) in healthy children aged 10–12 years. Nutr Neurosci. 2009; 12 (2): 48–56. 48. Stevens LJ, Zentall SS, Abate ML et al. Omega-3 fatty acids in boys with behavior, learning, and health problems. Physiol Behav. 1996; 59 (4–5): 915–920. 49. Stevens LJ, Zentall SS, Deck JL et al. Essential fatty acid metabolism in boys with attention-deficit hyperactivity disorder. Am J Clin Nutr. 1995; 62 (4): 761–768. 50. Antalis CJ, Stevens LJ, Campbell M, Pazdro R, Ericson K, Burgess JR. Omega-3 fatty acid status in attention-deficit/hyperactivity disorder. Prostaglandins Leukot Essent Fatty Acids. 2006; 75 (4–5): 299–308. 51. Chen JR, Hsu SF, Hsu CD, Hwang LH, Yang SC. Dietary patterns and blood fatty acid composition in children with attention-deficit hyperactivity disorder in Taiwan. J Nutr Biochem. 2004; 15 (8): 467–472. 52. Richardson AJ, Montgomery P. The Oxford-Durham study: a randomized, controlled trial of dietary supplementation with fatty acids in children with developmental coordination disorder. Pediatrics. 2005; 115 (5):1360–1366. 53. Richardson AJ, Puri BK. A randomized doubleblind, placebo-controlled study of the effects of supplementation with highly unsaturated fatty acids on ADHD-related symptoms in children with specific learning difficulties. Prog Neuropsychopharmacol Biol Psychiatry. 2002; 26 (2): 233–239. 54. Bloch MH, Qawasmi A. Omega-3 fatty acid supplementation for the treatment of children with attentiondeficit/hyperactivity disorder symptomatology: systematic review and meta-analysis. J Am Acad Child Adolesc Psychiatry. 2011; 50 (10): 991–1000. 55. Hirayama S, Hamazaki T, Terasawa K. Effect of docosahexaenoic acid-containing food administration on symptoms of attention-deficit/hyperactivity disorder – a placebo-controlled double-blind study. Eur J Clin Nutr. 2004; 58 (3): 467–473.
56. Schuchardt JP, Huss M, Stauss-Grabo M, Hahn A. Significance of long-chain polyunsaturated fatty acids (PUFAs) for the development and behaviour of children. Eur J Pediatr. 2010; 169 (2): 149–164. 57. D-A-CH (Hrsg.) DGE, ÖGE, SGE, SVE. Referenzwerte für die Nährstoffzufuhr. 2008. Neuer Umschau Buchverlag, Neustadt an der Weinstrasse. 58. Judge MP, Loosemore ED, DeMare CI, et al. Dietary docosahexaenoic acid (DHA) intake in pregnant women. J Am Diet Assoc. 2003; 103: A82. 59. Denomme J, Stark KD, Holub BJ. Directly quantitated dietary (n-3) fatty acid intakes of pregnant Canadian women are lower than current dietary recommendations. J Nutr 2005; 135 (2): 206–211. 60. DGE (Hrsg.) Ernährungsbericht. Deutsche Gesellschaft für Ernährung e. V. 2004, Bonn. 61. Garcia C, Millet V, Coste TC, Mimoun M, Ridet A, Antona C, Simeoni U, Armand M. French mothers’ milk deficient in DHA contains phospholipid species of potential interest for infant development. J Pediatr Gastroenterol Nutr. 2011; 53 (2): 206–212. 62. Brenna JT, Varamini B, Jensen RG, DiersenSchade DA, Boettcher JA, Arterburn LM. Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am J Clin Nutr. 2007; 85 (6): 1457–1664. 63. Koletzko B, Lien E, Agostoni C, Böhles H, Campoy C, Cetin I, Decsi T, Dudenhausen JW, Dupont C, Forsyth S, Hoesli I, Holzgreve W, Lapillonne A, Putet G, Secher NJ, Symonds M, Szajewska H, Willatts P, Uauy R; World Association of Perinatal Medicine Dietary Guidelines Working Group. The roles of long-chain polyunsaturated fatty acids in pregnancy, lactation and infancy: review of current knowledge and consensus recommendations. J Perinat Med. 2008; 36: 5–14. 64. Koletzko B, Cetin I, Brenna JT. Dietary fat intakes for pregnant and lactating women. Br J Nutr 2007; 98: 873–877. 65. Glaser C, Koletzko B. Langkettige Omega-3-Fettsäuren in der Perinatalzeit: Empfehlungen zur Zufuhr. Aktuel Ernährungsmed 2009; 34 (5): 240–245. 66. Koletzko B, Uauy R, Palou A, Kok F, Hornstra G, Eilander A, Moretti D, Osendarp S, Zock P, Innis S. Dietary intake of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) in children – a workshop report. Br J Nutr. 2010; 103 (6): 923–928.
2/12
12