


Transkript
ALKOHOL IN DER ERNÄHRUNG
Alkohol und Immunsystem
ALEXANDR PARLESAK
Der Konsum von Alkohol (hier: Ethanol) wirkt sich je nach Menge, Dauer der Einnahme, Art und Trinkmuster der alkoholischen Getränke sehr unterschiedlich auf die Immunantwort aus. Tendenziell betrachtet wirkt kurzfristiger Alkoholkonsum immunsupprimierend, während langfristiger Alkoholmissbrauch zu unspezifischen Entzündungsreaktionen führen kann, die auch für die alkoholbedingte Organschädigung von entscheidender Bedeutung sind. In jedem Fall führt langfristiger Alkoholmissbrauch zu einer eingeschränkten Resistenz gegenüber bakteriellen und viralen Infektionen, vor allem denen des Respirationstrakts. Hauptursachen hierfür sind eine eingeschränkte Phagozytosefähigkeit, eine Dysregulation der Bildung von immunrelevanten Botenstoffen (Zytokinen), eine gehemmte Antigenpräsentation sowie eine gestörte Chemotaxis. Eine wichtige Ursache der Dysregulation der Immunantwort durch chronischen Alkoholabusus liegt an einer erhöhten Permeabilität intestinaler bakterieller Toxine, die auf einer Einschränkung der intestinalen Barrierefunktion und einer bakteriellen Fehlbesiedlung des oberen Dünndarms beruht.
Vom Alkoholmissbrauch betroffene Bereiche der Immunantwort
Das Immunsystem muss die schwierige Aufgabe bewältigen, einerseits Toleranz gegenüber Millionen harmloser makromolekularer Strukturen zu entwickeln, die mit der Nahrung aufgenommen werden oder mit den Atemwegen in Kontakt kommen, und andererseits sehr schnell und effektiv Pathogene zu eliminieren. Diese Aufgabe erfüllt es sowohl mithilfe unspezifisch reagierender löslicher (humoraler) Faktoren, die Keime kurz vor oder nach dem Eindringen in den Körper abtöten (z.B. Defensine, Komplementfaktoren) als auch mithilfe spezifischer Antikörper, die wenige Tage nach Kontakt gegen das betreffende Pathogen gebildet werden. Weitverbreitete mikrobielle Strukturen (microbe-associated molecular patterns, MAMP) initiieren nach Erkennung durch zelluläre Rezeptoren (pattern recognition receptors, PRR) unterschiedli-
che unspezifische zelluläre Reaktionen (v.a. Phagozytose), die nach kurzer Zeit durch spezifische zelluläre Reaktionen (Expansion von T- und B-Zell-Klonen) sowie der Bildung von Antikörpern unterstützt werden. Diese einzelnen Bereiche sind über Signalmoleküle miteinander verbunden, sodass eine Störung der Immunantwort auch auf einer Dysregulation der Signalgebung durch diese Moleküle basieren kann. Jeder der einzelnen Bereiche der Immunantwort (humoral: unspezifisch, spezifisch; zellulär, unspezifisch und spezifisch) wird individuell in Abhängigkeit von Dauer und Art des Alkoholkonsums beeinflusst, wobei es zu einer erheblichen Interferenz des Alkohols mit der Kommunikation immunkompetenter Zellen untereinander kommt. Eine Übersicht über die Wirkungen des akuten und chronischen Alkoholmissbrauchs ist in der Tabelle wiedergegeben.
Konsequenzen des Alkoholmissbrauchs auf die Infektanfälligkeit
Obwohl gewissen Schwankungen unterworfen, ist bis zu einem täglichen Konsum von einem Drink (ca. 15 g Alkohol) für Frauen und zwei für Männer kein nachteiliger Effekt für das Immunsystem zu befürchten. Ein Drink entspricht (zumindest in der internationalen Fachliteratur) in etwa 0,35 Liter Bier beziehungsweise einem Glas (0,15 l) Wein oder 40 ml Schnaps. Voraussetzung hierfür ist, dass ein gleichförmiges Trinkmuster ohne Alkoholexzesse (wie z.B. Wochendbesäufnisse, «binge drinking») vorliegt. Von einem kritischen Umgang mit Alkohol (chronischer Alkoholmissbrauch) kann gesprochen werden, wenn Frauen mehr als zwei Drinks täglich (Männer: > 3,5 Drinks) über einen längeren Zeitraum (Monate oder Jahre) zu sich nehmen. Vor allem im Falle einer bereits bestehenden Schädigung der Leber (z.B. durch voran-
1/09
18
ALKOHOL IN DER ERNÄHRUNG
gehenden Alkoholmissbrauch, aber auch nach einer viralen Hepatitis) kann diese Toleranz jedoch herabgesetzt sein. Langfristiger erhöhter Alkoholkonsum ist ein Risikofaktor für unterschiedliche Infektionserkrankungen, vor allem der Lunge. In einer aktuellen Metaanalyse wurde eine Steigerung des relativen Risikos, an Tuberkulose zu erkranken, um den Faktor 2,9 bis 3,5 errechnet, wenn im Mittel mehr als 40 g Alkohol/Tag getrunken werden (1). Auch für bakterielle Pneumonie steigt das Risiko mit steigendem Alkoholkonsum (2), und Patienten mit dieser Erkrankung haben eine deutlich schlechtere Prognose bei vorangehendem Alkoholmissbrauch (3). Auch bei anderen Erkrankungen wie beispielsweise Cholera oder viraler Hepatitis sind die Erkrankungswahrscheinlichkeit und das Mortalitätsrisiko signifikant erhöht (4). Des Weiteren treten sowohl Sepsis als auch virale Hepatiden (z.B. Hepatitis C) nach chronischem Alkoholabusus häufiger auf, sind durch einen schwereren Krankheitsverlauf gekennzeichnet und haben eine deutlich schlechtere Prognose (5, 6). Obwohl der Zusammenhang einer erhöhten Erkrankungswahrscheinlichkeit für Infektionen einhergehend mit einem höheren Mortalitätsrisiko einerseits und exzessivem Alkoholgebrauch andererseits bereits 1785 von Benjamin Rush erkannt wurde, ist die Erkennung der Mechanismen, die Menschen mit exzessivem Alkoholkonsum zu einem immunsupprimierten Wirt machen, aufgrund der Komplexität der Immunantwort vielschichtig und noch nicht abgeschlossen. Die Analyse dieser Faktoren wird durch den Umstand erschwert, dass Menschen, die riskant mit Alkohol umgehen, auch ein in vielen Bereichen von der Norm abweichendes Sozialverhalten aufweisen (z.B. Rauchen, riskantes sexuelles Verhalten, ungeregelte Lebensweise, mangelnder Hygienestandard).
Auswirkungen des Alkoholmissbrauchs auf die unspezifische zelluläre Immunantwort
Der letztendlich zur Abtötung von Pathogenen führende Schritt ist die Phagozytose, bei der der Keim in ein Vesikel im
Phagozyten (Phagosom) eingeschlossen wird und mittels eines erniedrigten pH, der Freisetzung bakterizider Peptide, saurer Proteasen und reaktiver Sauerstoffverbindungen (reactive oxygen species, ROS) eliminiert wird. Neben der unmittelbaren Abtötung (v.a. durch neutrophile Granulozyten) dient die Phagozytose auch der Präsentation von Antigenen des Pathogens, die zur Aktivierung von B-Zellen und der Produktion von Antikörpern gegen Fragmente von Makromolekülen (Epitope) führt. Der letztgenannte Prozess wird vor allem durch dendritische Zellen und Makrophagen vorangetrieben. Bei mit Alkohol gefütterten Ratten weisen zwar Makrophagen aus Milz und dem Peritoneum signifikant höhere Konzentrationen phagozytoserelevanter Oberflächenrezeptoren (C3b und Fc) auf, sie sind aber dennoch nur zu einer verminderten Phagozytose von Candida-albicans-Zellen befähigt (7). Akute Alkoholexposition von Granulozyten und Monozyten des Menschen in vitro führt zu einer deutlichen Absenkung der Bildung von ROS nach Stimulation mit MAMP (8, 9). Bemerkenswerterweise ist der akute Effekt des Ethanols auf die ROS-Bildung – vermutlich adaptationsbedingt – bei Patienten mit moderater bis schwerer alkoholbedingter Schädigung der Leber nicht mehr nachweisbar (10). Auch andere Prozesse, die die Phagozytose unterstützen, sind bei Personen mit chronischem Alkoholmissbrauch gestört, vor allem dann, wenn die Leber durch Alkoholabusus bereits geschädigt ist (Alkoholhepatitis/-zirrhose). Hierzu gehören die Chemotaxis (Migration von Granulozyten zum Ort der bakteriellen Infektion) (11) und die Adhärenz von Granulozyten an das Endothel (12).
Wirkung des Alkohols auf das Zytokin- und Chemokinnetzwerk sowie andere immunrelevante Signalmoleküle
Klinisch relevante Alkoholkonzentrationen hemmen die Aktivierung immunkompetenter Zellen des Menschen ex vivo und in vitro durch den TLR2-Rezeptor (PRR für die Lipoteichonsäure, Peptidoglycan, Lipomannan und andere MAMP,
vor allem grampositiver Keime) praktisch nicht (13). Im Gegensatz dazu ist die Stimulation von Leukozyten über TLR3, einen Rezeptor, der für die Erkennung von (doppelsträngiger) RNA verantwortlich ist, bereits bei moderaten Alkoholkonzentrationen herabgesetzt (14). Dieser Umstand liefert eine mögliche Erklärung für die erhöhte Inzidenz viraler Erkrankungen bei Personen mit chronischem Alkoholmissbrauch. Analog hierzu ist die Erkennung der Oberflächenstrukturen gramnegativer Bakterien (Endotoxine = Lipopolysaccharide, LPS) durch Alkoholkonzentrationen, wie sie bei Alkoholikern auftreten, unterdrückt (15). Endotoxin ist ein klassisches MAMP und wird durch TLR4 mit den zugehörigen Korezeptoren CD14 und MD2) erkannt. Dieser Komplex startet eine Signalkaskade, die auch direkt durch proinflammatorische Zytokine (z.B. Interleukin 1-beta, IL-1β) stimuliert wird. Akute Alkoholexposition vermindert die Bildung dieser Zytokine, vor allem Tumor-Nekrose-Faktor alpha (TNF-α), IL-1β, IL-6 und anderer Zytokine, die von der Aktivierung des Transkriptionsfaktors NFκB abhängen oder die Signalwege ERK1/2 und MAPK nutzen (16–18). Dagegen werden Zytokine, die Entzündungen zum Abklingen bringen können (wie z.B. IL-10), unter Alkoholeinfluss vermehrt gebildet, basierend auf einer gesteigerten Aktivierung der Transkriptionsfaktoren STAT3 und AP-1 (19). Im Gegensatz dazu steigert chronischer Alkoholmissbrauch die Bildung inflammatorischer Zytokine durch immunkompetente Zellen (u. a. auch durch die kupfferschen Sternzellen), was zu einer erhöhten systemischen Konzentration von TNF-α, IL-1β, IL-6 und IL-8 bei Patienten mit alkoholinduzierter Lebererkrankung führt (20). Hierzu kommt es durch eine höhere Sensitivität ERK- und Egr1-assoziierter Signalwege (21), einer erhöhten Expression kostimulatorischer Moleküle (CD80/CD86) auf der Oberfläche immunkompetenter Zellen (22) und einer Stabilisierung der mRNA für einige dieser Zytokine (23), was zu einer weiteren Immunstimulation führt. Die erhöhte systemische Bildung von Chemokinen bei chronischer Alkoholexposition resultiert
19 1/09
ALKOHOL IN DER ERNÄHRUNG
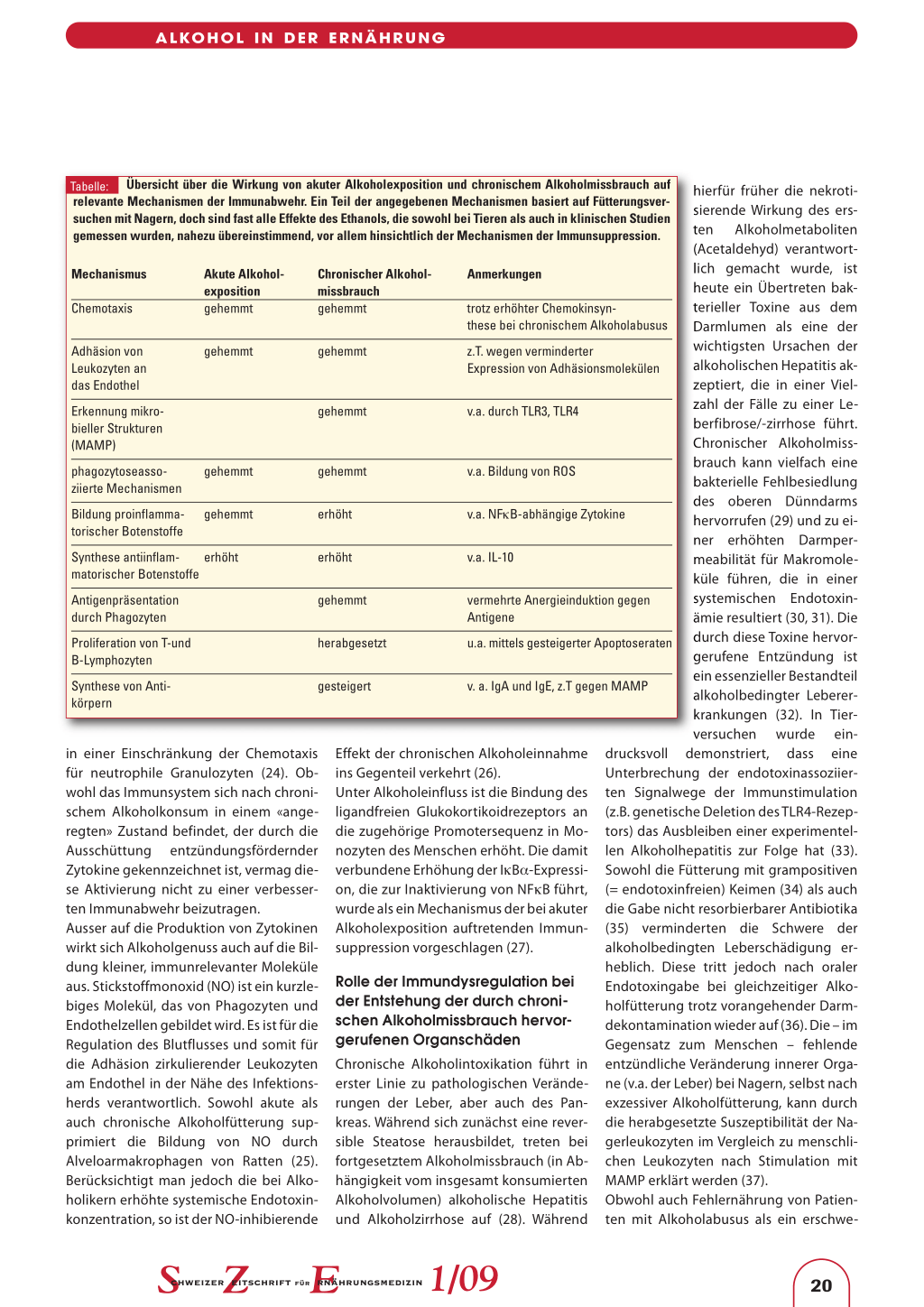
Tabelle: Übersicht über die Wirkung von akuter Alkoholexposition und chronischem Alkoholmissbrauch auf relevante Mechanismen der Immunabwehr. Ein Teil der angegebenen Mechanismen basiert auf Fütterungsversuchen mit Nagern, doch sind fast alle Effekte des Ethanols, die sowohl bei Tieren als auch in klinischen Studien gemessen wurden, nahezu übereinstimmend, vor allem hinsichtlich der Mechanismen der Immunsuppression.
hierfür früher die nekrotisierende Wirkung des ersten Alkoholmetaboliten (Acetaldehyd) verantwort-
Mechanismus Chemotaxis
Akute Alkoholexposition gehemmt
Chronischer Alkoholmissbrauch gehemmt
Anmerkungen trotz erhöhter Chemokinsyn-
lich gemacht wurde, ist heute ein Übertreten bakterieller Toxine aus dem
these bei chronischem Alkoholabusus Darmlumen als eine der
Adhäsion von Leukozyten an das Endothel
gehemmt
gehemmt
z.T. wegen verminderter Expression von Adhäsionsmolekülen
wichtigsten Ursachen der alkoholischen Hepatitis akzeptiert, die in einer Viel-
Erkennung mikrobieller Strukturen (MAMP)
gehemmt
v.a. durch TLR3, TLR4
zahl der Fälle zu einer Leberfibrose/-zirrhose führt. Chronischer Alkoholmiss-
phagozytoseassoziierte Mechanismen
Bildung proinflammatorischer Botenstoffe
gehemmt gehemmt
gehemmt erhöht
v.a. Bildung von ROS v.a. NFκB-abhängige Zytokine
brauch kann vielfach eine bakterielle Fehlbesiedlung des oberen Dünndarms hervorrufen (29) und zu einer erhöhten Darmper-
Synthese antiinflam- erhöht matorischer Botenstoffe
erhöht
v.a. IL-10
meabilität für Makromoleküle führen, die in einer
Antigenpräsentation
gehemmt
vermehrte Anergieinduktion gegen
systemischen Endotoxin-
durch Phagozyten
Antigene
ämie resultiert (30, 31). Die
Proliferation von T-und B-Lymphozyten
Synthese von Antikörpern
herabgesetzt gesteigert
u.a. mittels gesteigerter Apoptoseraten v. a. IgA und IgE, z.T gegen MAMP
durch diese Toxine hervorgerufene Entzündung ist ein essenzieller Bestandteil alkoholbedingter Lebererkrankungen (32). In Tier-
versuchen wurde ein-
in einer Einschränkung der Chemotaxis Effekt der chronischen Alkoholeinnahme drucksvoll demonstriert, dass eine
für neutrophile Granulozyten (24). Ob- ins Gegenteil verkehrt (26).
Unterbrechung der endotoxinassoziier-
wohl das Immunsystem sich nach chroni- Unter Alkoholeinfluss ist die Bindung des ten Signalwege der Immunstimulation
schem Alkoholkonsum in einem «ange- ligandfreien Glukokortikoidrezeptors an (z.B. genetische Deletion des TLR4-Rezep-
regten» Zustand befindet, der durch die die zugehörige Promotersequenz in Mo- tors) das Ausbleiben einer experimentel-
Ausschüttung entzündungsfördernder nozyten des Menschen erhöht. Die damit len Alkoholhepatitis zur Folge hat (33).
Zytokine gekennzeichnet ist, vermag die- verbundene Erhöhung der IκBα-Expressi- Sowohl die Fütterung mit grampositiven
se Aktivierung nicht zu einer verbesser- on, die zur Inaktivierung von NFκB führt, (= endotoxinfreien) Keimen (34) als auch
ten Immunabwehr beizutragen.
wurde als ein Mechanismus der bei akuter die Gabe nicht resorbierbarer Antibiotika
Ausser auf die Produktion von Zytokinen Alkoholexposition auftretenden Immun- (35) verminderten die Schwere der
wirkt sich Alkoholgenuss auch auf die Bil- suppression vorgeschlagen (27).
alkoholbedingten Leberschädigung er-
dung kleiner, immunrelevanter Moleküle aus. Stickstoffmonoxid (NO) ist ein kurzlebiges Molekül, das von Phagozyten und Endothelzellen gebildet wird. Es ist für die Regulation des Blutflusses und somit für
Rolle der Immundysregulation bei der Entstehung der durch chronischen Alkoholmissbrauch hervorgerufenen Organschäden
heblich. Diese tritt jedoch nach oraler Endotoxingabe bei gleichzeitiger Alkoholfütterung trotz vorangehender Darmdekontamination wieder auf (36). Die – im Gegensatz zum Menschen – fehlende
die Adhäsion zirkulierender Leukozyten Chronische Alkoholintoxikation führt in entzündliche Veränderung innerer Orga-
am Endothel in der Nähe des Infektions- erster Linie zu pathologischen Verände- ne (v.a. der Leber) bei Nagern, selbst nach
herds verantwortlich. Sowohl akute als rungen der Leber, aber auch des Pan- exzessiver Alkoholfütterung, kann durch
auch chronische Alkoholfütterung sup- kreas. Während sich zunächst eine rever- die herabgesetzte Suszeptibilität der Na-
primiert die Bildung von NO durch sible Steatose herausbildet, treten bei gerleukozyten im Vergleich zu menschli-
Alveloarmakrophagen von Ratten (25). fortgesetztem Alkoholmissbrauch (in Ab- chen Leukozyten nach Stimulation mit
Berücksichtigt man jedoch die bei Alko- hängigkeit vom insgesamt konsumierten MAMP erklärt werden (37).
holikern erhöhte systemische Endotoxin- Alkoholvolumen) alkoholische Hepatitis Obwohl auch Fehlernährung von Patien-
konzentration, so ist der NO-inhibierende und Alkoholzirrhose auf (28). Während ten mit Alkoholabusus als ein erschwe-
1/09
20
ALKOHOL IN DER ERNÄHRUNG
render Faktor sowohl für Infektionen als auch alkoholbedingte Lebererkrankungen diskutiert wurde (38), ist der mitteleuropäische Patient mit alkoholinduzierter Lebererkrankung nicht generell fehlernährt (39).
Auswirkungen des Alkoholmissbrauchs auf die spezifische humorale und zelluläre Immunantwort
Die klassische Präsentation von Antigenen durch Monozyten und durch dendritische Zellen des Menschen wird nach akuter Alkoholeinnahme gehemmt, was unter anderem durch eine Verschiebung des Verhältnisses antiinflammatorischer zu proinflammatorischen Zytokinen (IL10/IL-12) erklärt wurde (40, 41). Die adäquate Reaktion auf die Antigenpräsentation (in erster Linie die Produktion von Antikörpern) wird zusätzlich durch eine herabgesetzte Proliferation von T-Lymphozyten erschwert (41). Ein Zusatz exogener IL-12 kompensierte die durch Alkoholintoxikation hervorgerufene Anergie der T-Zellen. Erhöhte systemische Konzentrationen von Antikörpern der Klasse IgE sind bei Patienten mit Alkoholabusus weitverbreitet, ohne dass damit die Prävalenz für atopische Erkrankungen steigen würde (42). Auch die systemische Konzentration von IgA ist bereits bei moderaten Veränderungen der Leber durch chronischen Alkoholmissbrauch (Alkoholsteatose) erhöht, wobei ein Teil dieser Antikörperklasse gegen Endotoxine intestinalen Ursprungs gerichtet ist (43). Weitere antigene Strukturen für diese Antikörper beim Alkohol-missbrauchenden Patienten sind T-Zell-Proteine, DNA, Lipoproteine und unterschiedliche hepatische Proteine (44). Die Erhöhung dieser Konzentrationen basiert auf einer vermehrten Synthese der Immunglobuline, da die Anzahl zirkulierender B-Lymphozyten erniedrigt, das Verhältnis aktivierter (CD45RO+) zu naiven (CD45RA+) T-Zellen jedoch erhöht ist (45). Insgesamt tritt bei Alkoholikern allerdings eine Lymphopenie auf, die auf einer durch Ethanol bedingten Steigerung der Apoptoserate von T-Lymphozyten basiert (46).
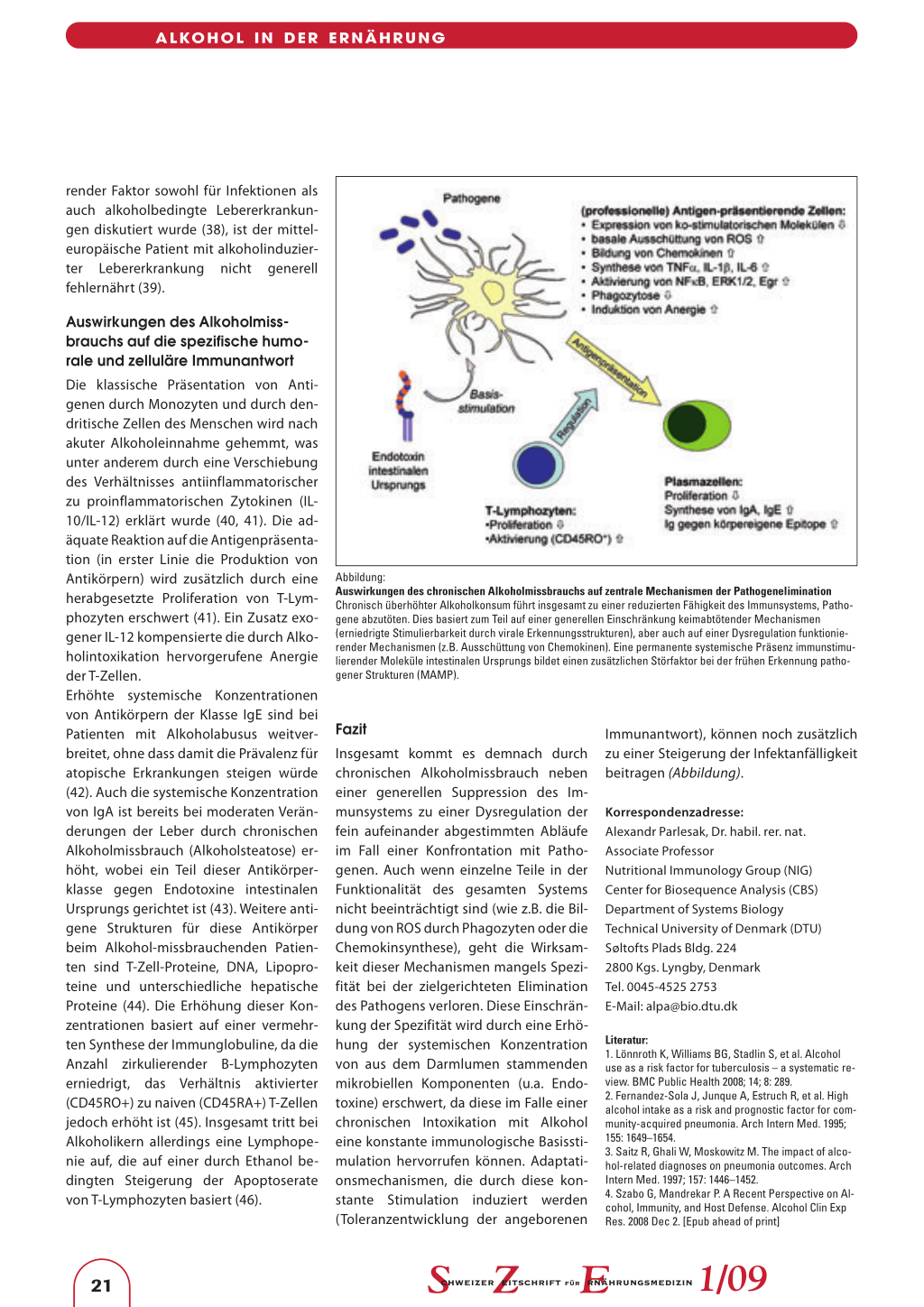
Abbildung: Auswirkungen des chronischen Alkoholmissbrauchs auf zentrale Mechanismen der Pathogenelimination Chronisch überhöhter Alkoholkonsum führt insgesamt zu einer reduzierten Fähigkeit des Immunsystems, Pathogene abzutöten. Dies basiert zum Teil auf einer generellen Einschränkung keimabtötender Mechanismen (erniedrigte Stimulierbarkeit durch virale Erkennungsstrukturen), aber auch auf einer Dysregulation funktionierender Mechanismen (z.B. Ausschüttung von Chemokinen). Eine permanente systemische Präsenz immunstimulierender Moleküle intestinalen Ursprungs bildet einen zusätzlichen Störfaktor bei der frühen Erkennung pathogener Strukturen (MAMP).
Fazit
Insgesamt kommt es demnach durch chronischen Alkoholmissbrauch neben einer generellen Suppression des Immunsystems zu einer Dysregulation der fein aufeinander abgestimmten Abläufe im Fall einer Konfrontation mit Pathogenen. Auch wenn einzelne Teile in der Funktionalität des gesamten Systems nicht beeinträchtigt sind (wie z.B. die Bildung von ROS durch Phagozyten oder die Chemokinsynthese), geht die Wirksamkeit dieser Mechanismen mangels Spezifität bei der zielgerichteten Elimination des Pathogens verloren. Diese Einschränkung der Spezifität wird durch eine Erhöhung der systemischen Konzentration von aus dem Darmlumen stammenden mikrobiellen Komponenten (u.a. Endotoxine) erschwert, da diese im Falle einer chronischen Intoxikation mit Alkohol eine konstante immunologische Basisstimulation hervorrufen können. Adaptationsmechanismen, die durch diese konstante Stimulation induziert werden (Toleranzentwicklung der angeborenen
Immunantwort), können noch zusätzlich zu einer Steigerung der Infektanfälligkeit beitragen (Abbildung).
Korrespondenzadresse: Alexandr Parlesak, Dr. habil. rer. nat. Associate Professor Nutritional Immunology Group (NIG) Center for Biosequence Analysis (CBS) Department of Systems Biology Technical University of Denmark (DTU) Søltofts Plads Bldg. 224 2800 Kgs. Lyngby, Denmark Tel. 0045-4525 2753 E-Mail: alpa@bio.dtu.dk
Literatur: 1. Lönnroth K, Williams BG, Stadlin S, et al. Alcohol use as a risk factor for tuberculosis – a systematic review. BMC Public Health 2008; 14; 8: 289. 2. Fernandez-Sola J, Junque A, Estruch R, et al. High alcohol intake as a risk and prognostic factor for community-acquired pneumonia. Arch Intern Med. 1995; 155: 1649–1654. 3. Saitz R, Ghali W, Moskowitz M. The impact of alcohol-related diagnoses on pneumonia outcomes. Arch Intern Med. 1997; 157: 1446–1452. 4. Szabo G, Mandrekar P. A Recent Perspective on Alcohol, Immunity, and Host Defense. Alcohol Clin Exp Res. 2008 Dec 2. [Epub ahead of print]
21 1/09
ALKOHOL IN DER ERNÄHRUNG
5. Moss M. Epidemiology of sepsis: race, sex, and chronic alcohol abuse. Clin Infect Dis. 2005; 41 Suppl 7: S490-S497. 6. McCullough AJ, O’Connor JF. Alcoholic liver disease: proposed recommendations for the American College of Gastroentrology. Am J Gastroenterol. 1998; 93: 2022–2036. 7. Bagasra O, Howeedy A, Kajdacsy-Balla A. Macrophage function in chronic experimental alcoholism. Immunology 1988; 65: 405–409. 8. Parlesak A, Diedrich JP, Schäfer C, Bode C. A low concentration of ethanol reduces the chemiluminescence of human granulocytes and monocytes but not the tumor necrosis factor alpha production by monocytes after endotoxin stimulation. Infect Immun. 1998; 66: 2809–2813. 9. Anthony V, Goodbey S, Hott J, Queener S. Alcoholinduced inhibition of alveolar macrophage oxidant releases in vivo and in vitro. Alcohol Clin Exp Res. 1993; 17: 389–393. 10. Parlesak A, Schäfer C, Paulus SB, et al. Phagocytosis and production of reactive oxygen species by peripheral blood phagocytes in patients with different stages of alcohol-induced liver disease: effect of acute exposure to low ethanol concentrations. Alcohol Clin Exp Res. 2003; 27: 503–508. 11. Fiuza C, Salcedo M, Clemente G, Tellado JM. In vivo neutrophil dysfunction in cirrhotic patients with advanced liver disease. J Infect Dis. 2000; 182: 526– 533. 12. Nilsson E, Lidstrom P, Patarroyo M, et al. Ethanol impairs certain aspects of neutrophil adhesion in vitro: comparisons with inhibition of expression of the CD18 antigen. J Infect Dis. 1991; 163: 591–597. 13. Dolganiuc A, Bakis G, Kodys K, et al. Acute ethanol treatment modulates Toll-like receptor-4 association with lipid rafts. Alcohol Clin Exp Res. 2006; 30: 76–85. 14. Pruett SB, Zheng Q, Fan R, et al. Ethanol suppresses cytokine responses induced through Toll-like receptors as well as innate resistance to Escherichia coli in a mouse model for binge drinking. Alcohol 2004; 33: 147–155. 15. Mandrekar P, Catalano D, Szabo G. Inhibition of lipopolysaccharide-mediated NFkappaB activation by ethanol in human monocytes. Int Immunol. 1999; 11: 1781–1190. 16. Pruett SB, Schwab C, Zheng Q, Fan R. Suppression of innate immunity by acute ethanol administration: a global perspective and a new mechanism beginning with inhibition of signaling through TLR3. J Immunol. 2004; 173: 2715–2724. 17. Mandrekar P, Jeliazkova V, Catalano D, Szabo G. Acute alcohol exposure exerts anti-inflammatory effects by inhibiting IκB kinase activity and p65 phosphorylation in human monocytes. J Immunol. 2007; 178: 7686–7693.
18. Goral J, Choudhry MA, Kovacs EJ. Acute ethanol exposure inhibits macrophage IL-6 production: role of p38 and ERK 1⁄2 MAPK. J Leukoc Biol. 2004; 75: 553– 559. 19. Norkina O, Dolganiuc A, Shapiro T, et al. Acute alcohol activates STAT3, AP-1, and Sp-1 transcription factors via the family of Src kinases to promote IL-10 production in human monocytes. J Leukoc Biol. 2007; 82: 752–762. 20. McClain C, Barve S, Deaciuc I, et al. Cytokines in alcoholic liver disease. Semin Liver Disease 1999; 19: 205–219. 21. Pritchard MT, Nagy LE. Ethanol-induced liver injury: potential roles for egr-1. Alcohol Clin Exp Res. 2005; 29: 146S–150S. 22. Lau AH, Abe M, Thomson AW. Ethanol affects the generation, cosignaling molecule expression, and function of plasmacytoid and myeloid dendritic cell subsets in vitro and in vivo. J Leukoc Biol 2006; 79: 911–953. 23. Nagy LE. Stabilization of tumor necrosis factor-alpha mRNA in macrophages in response to chronic ethanol exposure. Alcohol. 2004; 33: 229–233. 24. Bautista AP. Chronic alcohol intoxication primes Kupffer cells and endothelial cells for enhanced CCchemokine production and concomitantly suppresses phagocytosis and chemotaxis. Front Biosci. 2002; 7: a117–a125. 25. D’Souza NB, Nelson S, Summer WR, Deaciuc IV. Alcohol modulates alveolar macrophage tumor necrosis factor-alpha, superoxide anion, and nitric oxide secretion in the rat. Alcohol Clin Exp Res. 1996; 20: 156–163. 26. Enomoto N, Ikejima K, Kitamura T, et al. Alcohol enhances lipopolysaccharide-induced increases in nitric oxide production by Kupffer cells via mechanisms dependent on endotoxin. Alcohol Clin Exp Res. 2000; 24 (Suppl): 55S–58S. 27. Mandrekar P, Bellerose G, Szabo G. Inhibition of NF-kappa B binding correlates with increased nuclear glucocorticoid receptor levels in acute alcohol-treated human monocytes. Alcohol Clin Exp Res. 2002; 26: 1872–1879. 28. Lelbach WK. Cirrhosis in the alcoholic and its relation to the volume of alcohol abuse. Ann N Y Acad Sci. 1975; 252: 85–105. 29. Bode C, Bode JC. Effect of alcohol consumption on the gut. Best Pract Res Clin Gastroenterol. 2003; 17: 575–592. 30. Parlesak A, Schäfer C, Schütz T, et al. Increased intestinal permeability to macromolecules and endotoxemia in patients with chronic alcohol abuse in different stages of alcohol-induced liver disease. J Hepatol. 2000; 32: 742–747. 31. Fukui H, Brauner B, Bode JC, Bode C. Plasma endotoxin concentrations in patients with alcoholic and non-alcoholic liver disease: reevaluation with an im-
proved chromogenic assay. J Hepatol. 1991; 12: 162–169. 32. Nagy LE. Recent insights into the role of the innate immune system in the development of alcoholic liver disease. Exp Biol Med. 2003; 228: 882–889. 33. Uesugi T, Froh M, Arteel GE, et al. Toll-like receptor 4 is involved in the mechanism of early alcohol-induced liver injury in mice. Hepatology 2001; 34: 101–108. 34. Nanji AA, Khettry U, Sadrzadeh SM. Lactobacillus feeding reduces endotoxemia and severity of experimental alcoholic liver (disease). Proc Soc Exp Biol Med. 1994; 205: 243–247. 35. Adachi Y, Moore LE, Bradford BU, et al. Antibiotics prevent liver injury in rats following long-term exposure to ethanol. Gastroenterology 1995; 108: 218–224. 36. Mathurin P, Deng QG, Keshavarzian A, et al. Exacerbation of alcoholic liver injury by enteral endotoxin in rats. Hepatology 2000; 32: 1008–1017. 37. de la M Hall P, Lieber CS, DeCarli LM, et al. Models of alcoholic liver disease in rodents: a critical evaluation. Alcohol Clin Exp Res. 2001; 25 (5 Suppl): 254S–261S. 38. Lieber CS. Relationships between nutrition, alcohol use, and liver disease. Alcohol Res Health. 2003; 27: 220–231. 39. Bergheim I, Parlesak A, Dierks C, et al. Nutritional deficiencies in German middle-class male alcohol consumers: relation to dietary intake and severity of liver disease. Eur J Clin Nutr. 2003; 57: 431–438. 40. Szabo G, Mandrekar P, Catalano D. Inhibition of superantigen-induced T cell proliferation and monocyte IL-1b, TNF, and IL-6 production by acute ethanol treatment. J Leukoc Biol. 1995; 58: 342–350. 41. Mandrekar P, Catalano D, Dolganiuc A, et al. Inhibition of myeloid dendritic cell accessory cell function by alcohol correlates with reduced CD80⁄ CD86 expression and decreased IL-12 production. J Immunol. 2004; 173: 3398–3407. 42. Domínguez-Santalla MJ, Vidal C, Viñuela J, et al. Increased serum IgE in alcoholics: relationship with Th1/Th2 cytokine production by stimulated blood mononuclear cells. Alcohol Clin Exp Res. 2001; 25: 1198–1205. 43. Parlesak A, Schäfer C, Bode C. IgA against gut-derived endotoxins: does it contribute to suppression of hepatic inflammation in alcohol-induced liver disease? Dig Dis Sci. 2002; 47: 760–766. 44. Vidali M, Stewart SF, Albano E. Interplay between oxidative stress and immunity in the progression of alcohol-mediated liver injury. Trends Mol Med. 2008: 14: 63–71. 45. Cook RT, Waldschmidt TJ, Cook BL, et al. Loss of the CD5+ and CD45RAhi B cell subsets in alcoholics. Clin Exp Immunol. 1996; 103: 304–310. 46. Kapasi AA, Patel G, Goenka A, et al. Ethanol promotes T cell apoptosis through the mitochondrial pathway. Immunology 2003; 108: 313–320.
1/09
22