




Transkript
Neurobiologische Effekte von Alkohol
FORTBILDUNG
Für die Entwicklung einer Suchterkrankung sind gewisse krankheitsimmanente, neurobiologische Faktoren kennzeichnend. Im folgenden Beitrag werden insbesondere relevante Aspekte aus Molekulargenetik, Neurotransmission und funktioneller Bildgebung beschrieben und wird auf die Effekte von Alkohol auf die Emotionen fokussiert.
Lorenz Deutschenbaur Marc Walter
von Lorenz Deutschenbaur und Marc Walter
Einleitung
A lkohol zählt zu den meistgebrauchten psychoaktiven Substanzen weltweit, wobei die Alkoholabhängigkeit eines der schwerwiegendsten Probleme des öffentlichen Gesundheitssektors darstellt (1). Die Wirkung von Alkohol auf die Gefühlswelt wird als einer der wichtigsten Risikofaktoren für die Entwicklung einer Alkoholabhängigkeit angesehen (2). Das zentralnervöse System kann durch schädlichen und chronischen Alkoholkonsum verändert werden. Zu den bekannten Folgeerscheinungen zählen zum Beispiel der Rausch, delirante Zustandsbilder, Halluzinationen, die Wernicke-Enzephalopathie und demenzielle Entwicklungen. Doch auch vor und während der Entwicklung einer Suchterkrankung sind gewisse krankheitsimmanente, neurobiologische Faktoren kennzeichnend. Die Fokussierung auf biologische Aspekte erhebt nicht einen Hoheitsanspruch gegenüber psychodynamischen Vorgängen oder sozialen Faktoren, die bei der Erkrankung gleichsam Anteil haben. Auch soll ein Plädoyer für die neurobiologische Komponente der Abhängigkeitserkrankungen nicht in der Prä-Determinismus-Debatte subsumiert werden. Die pathophysiologischen Mechanismen der Alkoholabhängigkeit können allerdings zum Verständnis und zur Weiterentwicklung hochqualitativer und ressourcenorientierter Behandlungskonzepte im Sinne des biopsychosozialen Modells beitragen. Noch mehr kann ein für die Bevölkerung greifbares Krankheitsmodell zur Entstigmatisierung auf individueller Ebene und mehr Zugewandtheit im soziostrukturellen Kontext beitragen. Abhängiges Verhalten entsteht grundsätzlich aus der Verbindung genetischer (3) und umweltabhängiger Faktoren. Die genetischen Faktoren sind wirkungslos, wenn die Substanz mit Abhängigkeitspotenzial in einer Gesellschaft nicht verfügbar ist (4). Während Umweltfaktoren wie etwa Trinkgewohnheiten, soziale Akzeptanz des Konsums, Getränkekosten oder Verfügbarkeit über 70 Prozent des Auftretens eines Alkoholkonsums
erklären (5), wird die Entstehung einer Alkoholabhängigkeit zu zirka 50 bis 60 Prozent durch genetische Faktoren bestimmt (6). Darüber hinaus sind vor dem Hintergrund der hohen psychiatrischen Komorbiditätsraten bei Abhängigkeitserkrankungen auch mögliche gemeinsame genetische Grundlagen zu diskutieren (7). Befunde aus grossen epidemiologischen Untersuchungen legen nahe, dass das Muster dieser Komorbiditäten in Cluster externalisierender und internalisierender Störungen aufteilbar ist, wobei nach Zwillingsstudien gerade auch genetische Faktoren für das gleichzeitige Auftreten von Störungen beider Cluster verantwortlich zu sein scheinen (8). Der folgende Artikel soll neurobiologische Vulnerabilitätsfaktoren herausarbeiten, um die unterschiedlichen Effekte von Alkohol bei Gesunden und Kranken besser zu verstehen. Dabei werden insbesondere relevante Aspekte aus Molekulargenetik, Neurotransmission und funktioneller Bildgebung beschrieben. Hierbei wird primär auf die Effekte von Alkohol auf die Emotionen fokussiert.
Effekte von Alkohol auf die Neurotransmission Alkohol (Ethanol: C2H6O) beeinflusst viele Neurotransmittersysteme. Alkohol wird enzymatisch über die Alkohol-Dehydrogenase zu Acetaldehyd metabolisiert, woraus via Acetaldehyd-Dehydrogenase eine rasche Umwandlung in Acetat stattfindet. Acetaldehyd ist eine für den Organismus giftige Substanz, die für viele unerwünschte Wirkungen des Alkohols zuständig ist. Acetaldehyd bedingt generell die Denaturierung von Proteinstrukturen sowie die Stimulierung körpereigener Eiweisse (z.B. Zytokine) und führt dadurch zu den unliebsamen Vergiftungserscheinungen, die ab einer gewissen Alkoholmenge dem symptomatischen «Kater» zugrunde liegen. Alkohol entfaltet seine Wirkung nicht über einen einzelnen Rezeptortyp, sondern interagiert mit einer Vielzahl von Neurotransmittersystemen. Hierzu zählen vor allem opioiderge, serotonerge, gabaerge, glutamaterge sowie dopaminerge Transmissionssysteme. Die Aufzählung wichtiger Zusammenhänge der einzelnen Sys-
&4 1/2014
PSYCHIATRIE NEUROLOGIE
FORTBILDUNG
teme soll im Anschluss in ein neurobiologisches Interaktionsmodell einfliessen.
Das opioiderge Transmissionssystem Eine Interaktion besteht zwischen dem Endocannabinoidsystem und Alkohol (9). Die alkoholinduzierte Freisetzung endogener Opioide (z.B. Beta-Endorphin) hat an der Steuerung intrinsischer Belohnungssysteme Anteil. Diese Alkoholwirkung könnte bei Individuen besonders stark ausgeprägt sein, die eine erbliche Disposition zur Alkoholabhängigkeit zeigen. So zeigten Kinder alkoholabhängiger Patienten eine besonders starke Endorphinfreisetzung nach Alkoholkonsum (10). Auch im Tierversuch fand sich bei alkoholpräferierenden Tieren eine verstärkte Endorphinausschüttung nach Alkoholgabe. Generell ist die Endorphinausschüttung bei Alkoholikern eher niedrig, weswegen es zu einer kompensatorischen Zunahme postsynaptischer µ-Opioid-Rezeptoren kommt (11). Trifft nun eine alkoholinduzierte Endorphinfreisetzung auf postsynaptisch erhöhte µ-Opioid-Rezeptoren, kann es bei diesen disponierten Personen zu einer starken striären Dopaminfreisetzung kommen. In der Positronen-Emissions-Tomografie zeigte sich diesbezüglich eine deutliche Zunahme der µ-Opioid-Rezeptoren im ventralen Striatum alkoholabhängiger Patienten im Vergleich zu gesunden Kontrollpersonen. Die Erhöhung der µ-Opioid-Rezeptoren im ventralen Striatum und frontalen Kortex korrelierte dabei direkt mit dem Ausmass des chronischen Alkoholverlangens (12).
Das serotonerge Transmissionssystem Alkohol führt zu einer Erhöhung des Serotoninspiegels im zentralnervösen System, wobei verschiedene Studien für einen verminderten Serotoninumsatz bei alkoholabhängigen Patienten sprechen. Diese Beobachtung erscheint besonders relevant für eine Untergruppe alkoholabhängiger Patienten mit frühem Beginn der Alkoholabhängigkeit und «antisozialen» Persönlichkeitsmerkmalen (13). Denn ein verminderter Serotoninumsatz kann hierbei Grund für die «Eigenmedikation» durch Alkohol sein, wobei ein konditioniertes Lernen der disinhibierenden, anxiolytischen Wirkung denkbar ist. Ebenfalls kann ein primär insuffizientes serotonerges System Ausgangslage für eine entwicklungspsychologische soziale Fehlintegration sein. Nach chronischem Alkoholkonsum finden sich ebenfalls neurotoxisch bedingte Veränderungen, die primär mit negativen Gefühlszuständen wie Depressivität und Ängstlichkeit in Verbindung stehen (14). Die striäre Dopaminfreisetzung wird ebenfalls durch die serotonerge Neurotransmission moduliert. 5-HT3-Rezeptoren, die präsynaptisch auf dopaminergen Neuronen lokalisiert sind, stimulieren die Dopaminfreisetzung im Striatum. Die pharmakologische Blockade dieser Rezeptoren mittels Ondansetron führte bei Alkoholikern mit frühem Erkrankungsbeginn und sozialen Problemen zu einer Reduktion der Trinkmenge (15). In Kombination mit Naltrexon führte die spezifische 5-HT3Blockade zu einer verminderten Aktivierung des ventralen Striatums beim Zeigen alkoholassoziierter Bilder (16). Das kann als serotonerge Modulation einer für den Alkoholkonsum motivational wichtigen Region interpretiert werden.
Das gabaerge Transmissionssystem Alkohol aktiviert die inhibitorische Wirkung der GABAA-Rezeptoren, sodass ein übermässiger Alkoholkonsum subjektiv zur Sedation führt. Alkohol bindet hierbei direkt an eine aus Aminosäuren gebildete Tasche am GABA-A-Rezeptor und vermittelt so die Wirkung auf den Ionenkanal. Eine Toleranzentwicklung ist zum Teil auf eine reaktive Verminderung der GABA-A-Rezeptoren zurückzuführen, die sich im Entzug nur verzögert zurückbildet. Diese ist nach kurzfristiger Abstinenz noch nachweisbar (17). Bei plötzlicher Unterbrechung des Alkoholkonsums trifft der hirneigene Neurotransmitter GABA auf eine verminderte Rezeptorenzahl. Das Gleichgewicht zwischen Erregung und Dämpfung im Gehirn wird gestört, und die verminderte gabaerge Transmission trägt zur Entzugssymptomatik bei.
Das glutamaterge Transmissionssystem Kurz nach Einnahme von Alkohol wird die glutamaterge Transmission via NMDA-Rezeptor-Blockade generell herabgesetzt. Dabei lagert sich Ethanol an eine Bindungsstelle, über die sonst Glyzin die glutamaterge Übertragung verstärkt (18). Dementsprechend kommt es zu einer verminderten glutamatergen Neurotransmission am NMDA-Rezeptor, die direkt mit Gedächtnisstörungen bei exzessivem Alkoholkonsum in Verbindung stehen könnte (19). Bei chronischem Alkoholkonsum steigt nun zur Aufrechterhaltung der zentralnervösen Homöostase die Zahl und Funktion der NMDA-Rezeptoren reaktiv an. Entfällt die Alkoholzufuhr abrupt, verschiebt sich das zentralnervöse Gleichgewicht zwischen Exzitation und Sedation, und es kann zu Krampfanfällen kommen. Das akute Überwiegen der glutamatergen Exzitation im Entzug führt auch zur Enthemmung weiterer Botenstoffsysteme, wie beispielsweise zur Stimulation des Kerngebiets noradrenerger Nervenzellen im Locus couruleus des Hirnstamms, sodass es zum Auftreten weiterer, auch vegetativer Entzugssymptome kommen kann. Clonidin hemmt diese sekundär induzierte noradrenerge Transmission. In fortgeschrittenen Fällen genügt die Unterbrechung der Alkoholzufuhr während des Nachtschlafs, um eine morgendliche Entzugssymptomatik auszulösen. Neben den NMDA-Rezeptoren, die via Long-Term-Potentiation (LTP) kristalline Gedächtnisstrukturen und Plastizität im Gehirn modulieren, erfolgt der Einfluss auf das glutamaterge System ebenfalls mittels AMPARezeptoren (20), welche zu einer weiteren Depolarisierung an glutamatergen Synapsen führen. So wird zum Beispiel im Hippocampus die Dichte der AMPA-Rezeptoren in der postsynaptischen Membran abhängig von der Aktivität der Synapse reguliert (21). Diese Vorgänge spielen im Rahmen von Sensitivierungsprozessen eine wichtige Rolle. Eine Sensitivierung dopaminerger Zellverbände durch glutamaterge Long-Term-Potentiation könnte bei entsprechender Stimulation zu verstärkter Dopaminfreisetzung im Striatum führen und so einen wichtigen Faktor des Suchtgedächtnisses darstellen.
Das dopaminerge Transmissionssystem Gute Daten liegen für eine alkoholinduzierte Zunahme der dopaminergen Transmission im mesolimbischen System vor (22). Bei Laborratten stimuliert akuter wie chronischer Alkoholkonsum die Entladungsrate dop-
1/2014
PSYCHIATRIE & NEUROLOGIE
5
FORTBILDUNG
VTA DA neuron
VTA naltrexone
PFC glu neuron
DA enkephalin µ µ opioid receptor glu VSCC GABA NMDA receptor
GABAA receptor mGlu receptor
GABA Breceptor
µ
µ
naltrexone
GABA interneuron
arcuate nucleus opioid neuron
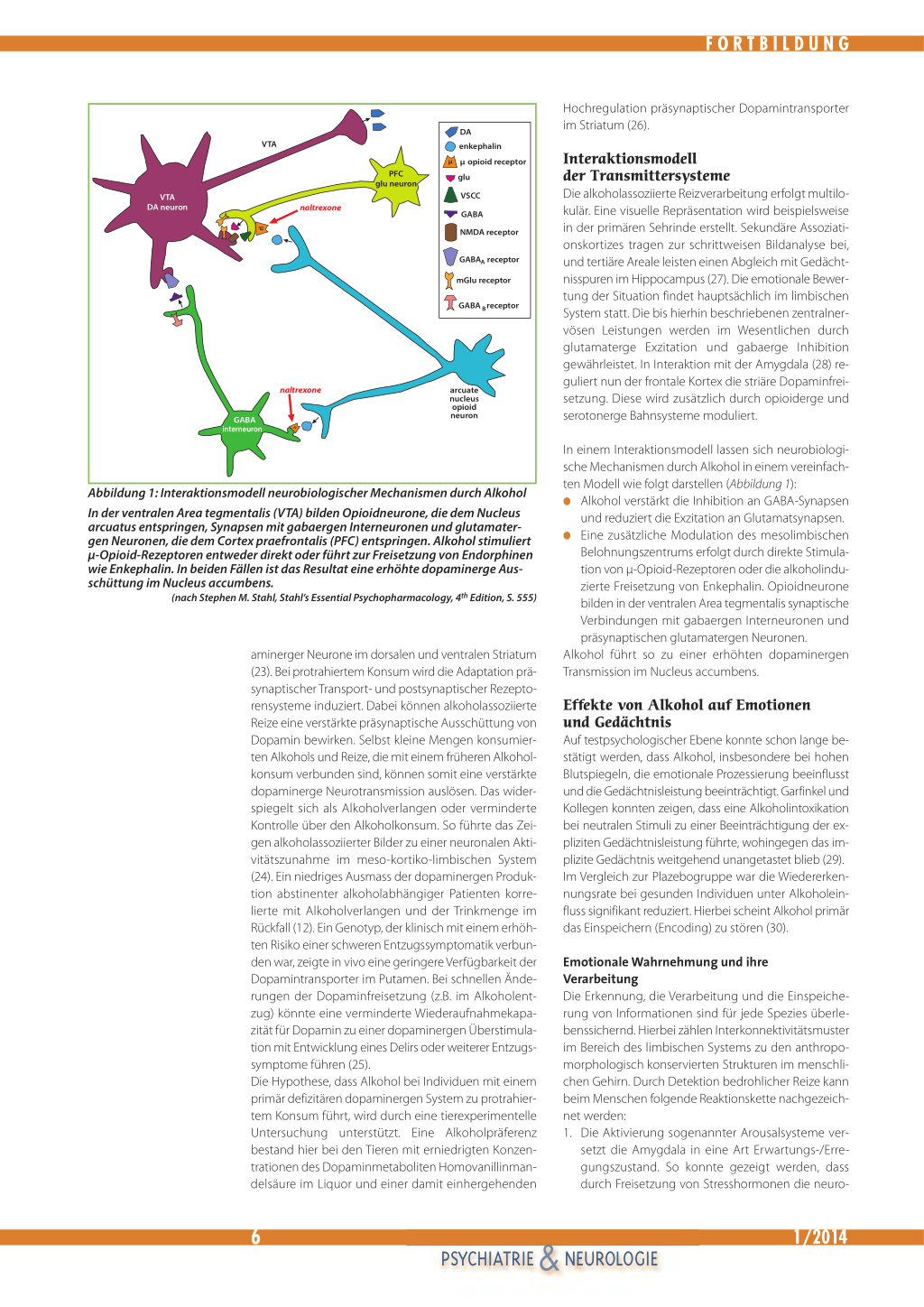
Abbildung 1: Interaktionsmodell neurobiologischer Mechanismen durch Alkohol
In der ventralen Area tegmentalis (VTA) bilden Opioidneurone, die dem Nucleus arcuatus entspringen, Synapsen mit gabaergen Interneuronen und glutamatergen Neuronen, die dem Cortex praefrontalis (PFC) entspringen. Alkohol stimuliert µ-Opioid-Rezeptoren entweder direkt oder führt zur Freisetzung von Endorphinen wie Enkephalin. In beiden Fällen ist das Resultat eine erhöhte dopaminerge Ausschüttung im Nucleus accumbens.
(nach Stephen M. Stahl, Stahl’s Essential Psychopharmacology, 4th Edition, S. 555)
aminerger Neurone im dorsalen und ventralen Striatum (23). Bei protrahiertem Konsum wird die Adaptation präsynaptischer Transport- und postsynaptischer Rezeptorensysteme induziert. Dabei können alkoholassoziierte Reize eine verstärkte präsynaptische Ausschüttung von Dopamin bewirken. Selbst kleine Mengen konsumierten Alkohols und Reize, die mit einem früheren Alkoholkonsum verbunden sind, können somit eine verstärkte dopaminerge Neurotransmission auslösen. Das widerspiegelt sich als Alkoholverlangen oder verminderte Kontrolle über den Alkoholkonsum. So führte das Zeigen alkoholassoziierter Bilder zu einer neuronalen Aktivitätszunahme im meso-kortiko-limbischen System (24). Ein niedriges Ausmass der dopaminergen Produktion abstinenter alkoholabhängiger Patienten korrelierte mit Alkoholverlangen und der Trinkmenge im Rückfall (12). Ein Genotyp, der klinisch mit einem erhöhten Risiko einer schweren Entzugssymptomatik verbunden war, zeigte in vivo eine geringere Verfügbarkeit der Dopamintransporter im Putamen. Bei schnellen Änderungen der Dopaminfreisetzung (z.B. im Alkoholentzug) könnte eine verminderte Wiederaufnahmekapazität für Dopamin zu einer dopaminergen Überstimulation mit Entwicklung eines Delirs oder weiterer Entzugssymptome führen (25). Die Hypothese, dass Alkohol bei Individuen mit einem primär defizitären dopaminergen System zu protrahiertem Konsum führt, wird durch eine tierexperimentelle Untersuchung unterstützt. Eine Alkoholpräferenz bestand hier bei den Tieren mit erniedrigten Konzentrationen des Dopaminmetaboliten Homovanillinmandelsäure im Liquor und einer damit einhergehenden
Hochregulation präsynaptischer Dopamintransporter im Striatum (26).
Interaktionsmodell der Transmittersysteme Die alkoholassoziierte Reizverarbeitung erfolgt multilokulär. Eine visuelle Repräsentation wird beispielsweise in der primären Sehrinde erstellt. Sekundäre Assoziationskortizes tragen zur schrittweisen Bildanalyse bei, und tertiäre Areale leisten einen Abgleich mit Gedächtnisspuren im Hippocampus (27). Die emotionale Bewertung der Situation findet hauptsächlich im limbischen System statt. Die bis hierhin beschriebenen zentralnervösen Leistungen werden im Wesentlichen durch glutamaterge Exzitation und gabaerge Inhibition gewährleistet. In Interaktion mit der Amygdala (28) reguliert nun der frontale Kortex die striäre Dopaminfreisetzung. Diese wird zusätzlich durch opioiderge und serotonerge Bahnsysteme moduliert.
In einem Interaktionsmodell lassen sich neurobiologische Mechanismen durch Alkohol in einem vereinfachten Modell wie folgt darstellen (Abbildung 1): G Alkohol verstärkt die Inhibition an GABA-Synapsen
und reduziert die Exzitation an Glutamatsynapsen. G Eine zusätzliche Modulation des mesolimbischen
Belohnungszentrums erfolgt durch direkte Stimulation von µ-Opioid-Rezeptoren oder die alkoholinduzierte Freisetzung von Enkephalin. Opioidneurone bilden in der ventralen Area tegmentalis synaptische Verbindungen mit gabaergen Interneuronen und präsynaptischen glutamatergen Neuronen. Alkohol führt so zu einer erhöhten dopaminergen Transmission im Nucleus accumbens.
Effekte von Alkohol auf Emotionen und Gedächtnis Auf testpsychologischer Ebene konnte schon lange bestätigt werden, dass Alkohol, insbesondere bei hohen Blutspiegeln, die emotionale Prozessierung beeinflusst und die Gedächtnisleistung beeinträchtigt. Garfinkel und Kollegen konnten zeigen, dass eine Alkoholintoxikation bei neutralen Stimuli zu einer Beeinträchtigung der expliziten Gedächtnisleistung führte, wohingegen das implizite Gedächtnis weitgehend unangetastet blieb (29). Im Vergleich zur Plazebogruppe war die Wiedererkennungsrate bei gesunden Individuen unter Alkoholeinfluss signifikant reduziert. Hierbei scheint Alkohol primär das Einspeichern (Encoding) zu stören (30).
Emotionale Wahrnehmung und ihre Verarbeitung Die Erkennung, die Verarbeitung und die Einspeicherung von Informationen sind für jede Spezies überlebenssichernd. Hierbei zählen Interkonnektivitätsmuster im Bereich des limbischen Systems zu den anthropomorphologisch konservierten Strukturen im menschlichen Gehirn. Durch Detektion bedrohlicher Reize kann beim Menschen folgende Reaktionskette nachgezeichnet werden: 1. Die Aktivierung sogenannter Arousalsysteme ver-
setzt die Amygdala in eine Art Erwartungs-/Erregungszustand. So konnte gezeigt werden, dass durch Freisetzung von Stresshormonen die neuro-
&6 1/2014
PSYCHIATRIE NEUROLOGIE
FORTBILDUNG
nale Aktivität (firing) im basolateralen Teil der Amygdala zunahm (31). 2. Die Amygdala stellt dem Hypothalamus zentralnervöse Repräsentanzen bedrohlich erkannter Faktoren zur Verfügung. 3. Im Rahmen der Hypothalamus-HypophysenNebennierenrinden-(HHN)-Achse erfolgt via Hypophyse die Freisetzung des adrenocorticotropen Hormons, welches auf peripherer Drüsenebene zur Ausschüttung von Cortisol und weiterer Stresshormone führt. Cortisol wiederum passiert die BlutHirn-Schranke und bindet an die entsprechenden Rezeptoren, unter anderem im Hippocampus, in der Amygdala und im präfrontalen Kortex (32). Bei gesunden Individuen kann Alkohol – ähnlich anderer psychoaktiver Substanzen wie Cannabis oder Heroin – mehrere positive und negative Effekte auf die Emotionsverarbeitung und das emotionale Erleben entfalten. Tierversuche zeigen, dass Alkohol zu einer Reduktion negativer Emotionen führt und Muster ängstlichen Verhaltens abdämpft (33). Selbsterfahrungsberichte mit Alkoholkonsum umfassen Euphorie, Entspannung, Enthemmung sowie Stress- und Angstreduktion. Die Alkoholintoxikation wird mit emotionaler Labilität und einer gestörten kognitiven Kontrollfunktion in Verbindung gebracht (34).
Emotionales Gedächtnis Bedrohliche Umweltsignale führen neben den viel zitierten «fight or flight»-Reaktionen, die via autonomes Nervensystem koordiniert werden, ebenso zu emotionalen Verarbeitungsprozessen, die durch Strukturen des limbischen Systems aufbereitet und in folgliche Handlungsmuster eingespeist werden (Abbildung 2). Dass die Amygdala eine Schlüsselrolle beim Prozessieren und bei der Einspeicherung emotionaler Sinneseindrücke spielt, beweisen zahlreiche Untersuchungen an Tier und Mensch: Die Amygdalaaktivität wird durch die Erkennung emotional wahrgenommener Sinnesreize (emotionales Arousal) stimuliert. Der Grad der Aktivierung wiederum korreliert eng mit der Einspeicherung und somit der Fähigkeit, emotionale Gedächtnisinhalte später wieder abrufen zu können (35). Insbesondere der basolaterale Teil der Amygdala, der eine hohe Konnektivität mit dem Hippocampus aufweist, fungiert als eine Art Booster in der Einspeicherung emotional gefärbter Inhalte (35). Das spiegelt den Eindruck wider, dass emotionale Informationsinhalte generell besser erinnert werden können.
Alkohol und emotionales Gedächtnis Eine Alkoholabhängigkeit manifestiert sich häufig um das 25. Lebensjahr. Neben einer Reihe weiterer Faktoren (männliches Geschlecht, Singlestatus) spielt das «Ansprechen» (Auslösen alkoholassoziierter affektiver und kognitiver Zustandsänderungen durch eine standardisierte Trinkmenge) auf Alkohol eine wichtige Rolle bei der Entwicklung einer späteren Suchterkrankung (36). Alkoholkonsum führt bei gesunden Individuen unter anderem zu einer Veränderung des Cortisolspiegels und damit einhergehend zu einer verminderten Stresswahrnehmung. Eine veränderte Amygdalafunktion konnte in diesem Zusammenhang bei Gesellschaftstrinkern, genau wie
bei Angsterkrankungen und sozialen Phobien, gezeigt werden. Bei schweren Alkoholikern bestand diese Korrelation nicht (37). Bei Studenten mit hohem Risiko eines Alkoholabusus konnte eine Amygdalaaktivierung lediglich durch Zeigen von Bildern mit negativen Emotionen verzeichnet werden. Das Zeigen von Bildern mit positiven Emotionen oder das Zeigen von Alkoholutensilien war hauptsächlich mit einer Aktivierung präfrontaler Hirnregionen assoziiert (38). Diese Ergebnisse stützen die Hypothese, dass die anxiolytischen und stressreduzierenden Faktoren des Alkoholkonsums durch eine Herabsetzung der Amygdalareaktivität vermittelt werden. Insbesondere junge Erwachsene mit hohem Risikoprofil führen den Alkoholkonsum fort, wenn sie die Erfahrung einer durch Alkohol veränderten zentralnervösen Reaktivität gemacht haben. Studien, die Gedächtnisleistung in Bezug auf emotionale Stimuli untersuchten, erbrachten im Zusammenhang mit Alkohol folgende Ergebnisse:
1. Encoding: Bei einer Untersuchung des impliziten Gedächtnisses mit repetitivem «Priming Task» (individuelle Reaktion auf die Präsentation eines erstmals gezeigten Stimulus zu einem späteren Zeitpunkt) zeichnete sich ab, dass im Rahmen einer Alkoholintoxikation das Priming für positive und negative emotionale Stimuli besser ausfiel im Vergleich zu neutralen Stimuli (30).
2. Recall: Im Rahmen einer Alkoholintoxikation war der Abruf neutraler wie auch emotionaler Stimuli signifikant erschwert (30).
3. Reaktivität: Alkohol setzte die Reaktion auf negativ emotionale Stimuli in der limbischen Region herab: Das Zeigen ängstlicher sowie wütender Gesichtsausdrücke erzeugte eine herabgesetzte AmygdalaAntwort unter Alkoholeinfluss (39).
Der modulierende Effekt von Alkohol auf die Verarbeitung emotionaler Stimuli und deren Gedächtniskonsolidierung könnte in Zusammenhang mit einer «Selbstmedikation» bei vulnerablen Individuen stehen. Vulnerabilität bezieht sich hier auf zentralnervöse Alterationen, die durch Alkohol zu einer subjektiv verbesserten Reaktivität/Verarbeitungsleistung führen und so den protrahierten Konsum bahnen.
Effekte von Alkohol auf Verhalten und psychiatrische Komorbidität Verhaltensebene der Sucht Eine Abhängigkeitsentwicklung geht mit einer Schwäche oder einer Störung der Verhaltensplanung und der Verhaltenskontrolle in Bezug auf die Bewertung langfristiger Ziele einher, wohingegen die positiven Wirkungen des Suchtmittelkonsums, die kurzfristig erreichbar sind, entsprechende Handlungsentwürfe bahnen (40). Der Iterationsprozess vom Handlungsentwurf bis zu dessen ausgewogener Umsetzung (Verhaltenskontrolle) findet eine Repräsentanz im dorsolateralen, präfrontalen Kortex, wobei das anteriore Cingulum bei konkurrierenden Handlungsmöglichkeiten (pro und kontra Alkoholkonsum) zusätzlich eingeschaltet wird (41). Eine Störung des frontalen Kortex kann bei verschiedenen Erkrankungen (z.B. alkoholbedingte Schädigung des Fetus, Hirntraumen, Disposition zum ADHS)
&8 1/2014
PSYCHIATRIE NEUROLOGIE
FORTBILDUNG
auftreten. Die neuronalen Strukturen des frontalen Kortex wiederum, die der Generierung kontrollierter Handlungen zugrunde liegen, werden durch chronischen Alkoholkonsum besonders beeinträchtigt (42). Gerade bei Patienten mit langer Dauer der Abhängigkeitserkrankung löst die Droge oft kaum noch angenehme Gefühle aus. Trotzdem denken diese Patienten oft obsessiv an den Drogenkonsum (43). Die neurobiologischen Strukturen (orbitofrontaler Kortex, Caput caudati), die diesem zwanghaften Alkoholverlangen zugrunde liegen können, sind dieselben, die auch bei der Entstehung anankastischen Verhaltens bei Patienten mit einer Zwangsstörung beteiligt sein sollen (44). Im hypothetischen Modell kommt es zur Generierung stereotyper Handlungsschablonen auf Basalganglienebene. Diese werden durch zentralnervöse Verbände wie dem orbitofrontalen Kortex nicht ausreichend gefiltert und entfalten Handlungsrelevanz. Eine Handlungskontrolle durch Integration emotionaler oder rationaler Repräsentanzen via dorsolateralen und präfrontalen Kortex entfällt bei dessen Dysfunktion.
Psychiatrische Komorbidität Ein bekanntes Beispiel für die Eigenmedikation einer affektiv bipolaren Erkrankung ist der Maler Vincent van Gogh, der seinen Umgang mit der Erkrankung folgendermassen beschrieb: «If the storm within gets too loud, I take a glass too much to stun myself» (45). «Wenn der Sturm in mir zu sehr tobt, nehme ich ein Glas zu viel, um mich zu betäuben.» Auch heutzutage noch behandeln viele Menschen eine psychiatrische Erkrankung mit Alkohol. Epidemiologische Feldstudien belegen ein 2,2-fach erhöhtes Risiko für Angsterkrankungen bei Individuen mit Alkoholabhängigkeit im Vergleich zur Allgemeinbevölkerung (46), wobei soziale und spezifische Phobien die häufigsten sind (47). Alkoholkonsum geht mit einer Angstreduktion und einer Störung der Gedächtnisfunktion einher. Die Selbstmedikation von Angsterkrankungen mit Alkohol kann partiell die hohe Komorbidität erklären, wobei ebenfalls die eingangs erwähnte genetische Clusterverteilung eine Rolle spielt. Im Vergleich zur Allgemeinbevölkerung zeigen Individuen mit einer Alkoholabhängigkeit eine zweifach erhöhte Prävalenz depressiver Störungen. Die durchschnittliche Lebenszeitprävalenz für eine Alkoholerkrankung war bei Patienten mit einer depressiven Störung deutlich erhöht (16–30% im Vergleich zur Allgemeinbevölkerung mit 7–16% [46]). Für viele dieser Störungen finden sich gemeinsame Risikofaktoren sowie im Verlauf der Erkrankungen ähnliche neurobiologische Veränderungen. In der Behandlung sollte eine individuelle, in den meisten Fällen kombinierte pharmakologische und psychosoziale Intervention erfolgen (siehe Artikel Seite 16 und 23). Eine parallele und integrierte Behandlung der komorbiden Störungsbilder ist der Goldstandard, um den häufig vorliegenden «Teufelskreis» zu durchbrechen und um eine positive Entwicklung einzuleiten (48).
Ausblick Alkohol verändert die Neurotransmission, führt zu Veränderungen in der emotionalen Prozessierung und der Gedächtnisfunktion. Bei vulnerablen Personen können
Unkonditionierte Angst olfaktorische Stimuli
akustische Stimuli
vomeronasales System auditorisch-visuelles System
sensorische Systeme
Konditionierte Angst visuelle, auditorische und olfaktorische Stimuli
auditorisches/visuelles/olfaktorisches System
LA MEA ABA
VMH PMH
Amygdala Hypothalamus
LA BA ABA ITC
CEA
PAGd
Dienzephalon
PAGv
– «Fight and Flight» – Wut, Aggressivität,
Enttäuschung – dissoziale Muster
motorische, emotionale, präfrontale
Systeme
– «Fight and Flight» – Wut, Aggressivität,
Enttäuschung – dissoziale Muster
Abbildung 2: Kreislauf der Abwehrreaktionen aufgrund konditionierter und unkonditionierter Ängste
Abkürzungen: ABA: akzessorisch-(basale) Amygdala; BA: basale Amygdala; CEA: zentrale Amygdala; LA: laterale Amygdala; LH: lateraler Hypothalamus; MEA: mediale Amygdala; NAcc: Nucleus accumbens; VMH: ventromedialer Hypothalamus; PAGd: dorsales periaquäduktales Grau; PAGv: ventrales periaquäduktales Grau; PMH: prämammilärer Hypothalamus
(adaptiert nach: Le Doux 2012)
diese Faktoren zu einer Suchtentwicklung beitragen.
Vulnerabilität bezieht sich hier auf Individuen mit krank-
heitsimmanenten Veränderungen. Anhand der bis dato
zur Verfügung stehenden wissenschaftlichen Datenlage
zeichnet sich ab, dass diese Veränderungen (z.B. des lim-
bischen Systems) schon vorbestehen. Beim Kontakt mit
einer Substanz, die gemäss den Kriterien abhängig ma-
chen kann (wie hier Alkohol), trägt die entsprechende
Prädisposition im Rahmen des individuellen psychody-
namischen und soziokulturellen Umfelds zur Krank-
heitsentwicklung und Aufrechterhaltung der Sucht bei.
Weitere Forschung ist notwendig, um mehr über die In-
teraktion von Alkohol, emotionaler Verarbeitung und
Suchtentwicklung zu erfahren. Inwieweit könnte zum
Beispiel eine primäre Veränderung des limbischen
Systems und hier insbesondere der Amygdala zur Ent-
wicklung einer Alkoholerkrankung disponieren? Wie
könnten Betroffene früher erkannt werden und Zugang
zu professioneller Hilfe erhalten? Zielgerichtete Thera-
pien könnten sich an weiteren Erkenntnissen aus
diesem Bereich orientieren, wobei ein Behandlungskon-
zept immer auch in ein biopsychosoziales Modell ein-
gebettet sein sollte.
G
Korrespondenzadresse:
Dr. med. Lorenz Deutschenbaur
Erwachsenen-Psychiatrische Klinik
Universitäre Psychiatrische Kliniken Basel
Wilhelm Klein-Strasse 27
4025 Basel
E-Mail: lorenz.deutschenbaur@upkbs.ch
Tel. 061-325 53 04
1/2014
PSYCHIATRIE & NEUROLOGIE
9
FORTBILDUNG
Fazit:
G Die Wirkung von Alkohol auf die Gefühlswelt
wird als einer der wichtigsten Risikofaktoren
für die Entwicklung einer Alkoholabhängigkeit
angesehen.
G Abhängiges Verhalten entsteht grundsätzlich
aus der Verbindung genetischer und umwelt-
abhängiger Faktoren.
G Alkohol entfaltet seine Wirkung nicht über
einen einzelnen Rezeptortyp, sondern inter-
agiert mit einer Vielzahl von Neurotransmitter-
systemen. Hierzu zählen vor allem opioiderge,
serotonerge, gabaerge, glutamaterge sowie
dopaminerge Transmissionssysteme.
G Alkohol verändert die Neurotransmission,
führt zu Veränderungen in der emotionalen
Prozessierung und der Gedächtnisfunktion. Bei
vulnerablen Personen können diese Faktoren
zu einer Suchtentwicklung beitragen.
Literatur: 1. Barnes, D. M.: Drugs: running the numbers. Science 240, 1729–1731
(1988). 2. Ray, L. A., MacKillop, J., Leventhal, A., Hutchison, K. E.: Catching the
alcohol buzz: an examination of the latent factor structure of subjective intoxication. Alcohol Clin Exp Res 33, 2154–2161, doi:10.1111/j.1530-0277.2009.01053.x (2009). 3. Kendler, K. S., Karkowski, L. M., Neale, M. C., Prescott, C. A.: Illicit psychoactive substance use, heavy use, abuse, and dependence in a US population-based sample of male twins. Archives of general psychiatry 57, 261–269 (2000). 4. Mann, K., Hermann, D., Heinz, A.: One hundred years of alcoholism: the Twentieth Century. Alcohol and alcoholism 35, 10–15 (2000). 5. Maes, H. H. et al.: Tobacco, alcohol and drug use in eight- to sixteenyear-old twins: the Virginia Twin Study of Adolescent Behavioral Development. Journal of studies on alcohol 60, 293–305 (1999). 6. True, W. R. et al.: Common genetic vulnerability for nicotine and alcohol dependence in men. Archives of general psychiatry 56, 655–661 (1999). 7. Compton, W. M., Thomas, Y. F., Stinson, F. S., Grant, B. F.: Prevalence, correlates, disability, and comorbidity of DSM-IV drug abuse and dependence in the United States: results from the national epidemiologic survey on alcohol and related conditions. Archives of general psychiatry 64, 566-576, doi:10.1001/archpsyc.64.5.566 (2007). 8. Kendler, K. S., Prescott, C. A., Myers, J., Neale, M. C.: The structure of genetic and environmental risk factors for common psychiatric and substance use disorders in men and women. Archives of general psychiatry 60, 929–937, doi:10.1001/archpsyc.60.9.929 (2003). 9. Economidou, D. et al.: Effect of the cannabinoid CB1 receptor antagonist SR-141716A on ethanol self-administration and ethanol-seeking behaviour in rats. Psychopharmacology 183, 394–403, doi:10.1007/s00213-005-0199-9 (2006). 10. Froehlich, J. C., Zink, R. W., Li, T. K., Christian, J. C.: Analysis of heritability of hormonal responses to alcohol in twins: beta-endorphin as a potential biomarker of genetic risk for alcoholism. Alcohol Clin Exp Res 24, 265–277 (2000). 11. Cowen, M. S., Lawrence, A. J.: The role of opioid-dopamine interactions in the induction and maintenance of ethanol consumption. Progress in neuro-psychopharmacology & biological psychiatry 23, 1171–1212 (1999). 12. Heinz, A. et al.: Correlation of stable elevations in striatal mu-opioid receptor availability in detoxified alcoholic patients with alcohol craving: a positron emission tomography study using carbon 11-labeled carfentanil. Archives of general psychiatry 62, 57–64, doi:10.1001/ archpsyc.62.1.57 (2005). 13. Fils-Aime, M. L. et al.: Early-onset alcoholics have lower cerebrospinal fluid 5-hydroxyindoleacetic acid levels than late-onset alcoholics. Archives of general psychiatry 53, 211–216 (1996). 14. Heinz, A. et al.: In vivo association between alcohol intoxication, aggression, and serotonin transporter availability in nonhuman primates. Am J Psychiatry 155, 1023–1028 (1998). 15. Ait-Daoud, N., Johnson, B. A., Prihoda, T. J., Hargita, I. D.: Combining ondansetron and naltrexone reduces craving among biologically predisposed alcoholics: preliminary clinical evidence. Psychopharmacology (Berl) 154, 23–27 (2001). 16. Myrick, H. et al.: Effect of naltrexone and ondansetron on alcohol cueinduced activation of the ventral striatum in alcohol-dependent people. Archives of general psychiatry 65, 466–475, doi:10.1001/archpsyc.65.4.466 (2008). 17. Abi-Dargham, A. et al.: Alterations of benzodiazepine receptors in type II alcoholic subjects measured with SPECT and [123I]iomazenil. Am J Psychiatry 155, 1550–1555 (1998).
18. Mascia, M. P., Trudell, J. R., Harris, R. A.: Specific binding sites for alcohols and anesthetics on ligand-gated ion channels. Proc Natl Acad Sci U S A 97, 9305–9310, doi:10.1073/pnas.160128797 (2000).
19. Tsai, G., Gastfriend, D. R., Coyle, J. T.: The glutamatergic basis of human alcoholism. Am J Psychiatry 152, 332–340 (1995).
20. Krystal JH, T. B. I. D. & K, C. D., Coyle JT, Nemeroff C, editors: Philadelphia, PA. Ethanol abuse, dependence, and withdrawal: neurobiology and clinical implications. Lippincott Williams & Wilkins, pp 1425–1443 (2002).
21. Huganir, R. L., Nicoll, R. A.: AMPARs and Synaptic Plasticity: The Last 25 Years. Neuron 80, 704–717, doi:10.1016/j.neuron.2013.10.025 (2013).
22. Johnson, B. A.: Update on neuropharmacological treatments for alcoholism: scientific basis and clinical findings. Biochemical pharmacology 75, 34–56, doi:10.1016/j.bcp.2007.08.005 (2008).
23. Imperato, A., Di Chiara, G.: Preferential stimulation of dopamine release in the nucleus accumbens of freely moving rats by ethanol. J Pharmacol Exp Ther 239, 219–228 (1986).
24. Buhler, M., Mann, K.: Alcohol and the human brain: a systematic review of different neuroimaging methods. Alcoholism, clinical and experimental research 35, 1771–1793, doi:10.1111/j.1530-0277.2011. 01540.x (2011).
25. Sander, T. et al.: Serotonin transporter gene variants in alcohol-dependent subjects with dissocial personality disorder. Biol Psychiatry 43, 908–912 (1998).
26. Mash, D. C. et al.: Altered dopamine transporter densities in alcoholpreferring vervet monkeys. Neuroreport 7, 457–462 (1996).
27. Vorel, S. R., Liu, X., Hayes, R. J., Spector, J. A., Gardner, E. L.: Relapse to cocaine-seeking after hippocampal theta burst stimulation. Science 292, 1175-1178, doi:10.1126/science.1058043 (2001).
28. Jackson, M. E., Moghaddam, B.: Amygdala regulation of nucleus accumbens dopamine output is governed by the prefrontal cortex. J Neurosci 21, 676–681 (2001).
29. Garfinkel, S. N., Dienes, Z., Duka, T.: The effect of alcohol and repetition at encoding on implicit and explicit false memories. Psychopharmacology 188, 498–508, doi:10.1007/s00213-006-0480-6 (2006).
30. Ray, S., Bates, M. E.: Acute alcohol effects on repetition priming and word recognition memory with equivalent memory cues. Brain and cognition 60, 118-127, doi:10.1016/j.bandc.2005.07.009 (2006).
31. Pelletier, J. G., Pare, D.: Role of amygdala oscillations in the consolidation of emotional memories. Biol Psychiatry 55, 559–562, doi:10.1016/j.biopsych.2003.08.019 (2004).
32. LeDoux, J.: The emotional brain, fear, and the amygdala. Cellular and molecular neurobiology 23, 727–738 (2003).
33. Spanagel, R. et al.: Anxiety: a potential predictor of vulnerability to the initiation of ethanol self-administration in rats. Psychopharmacology 122, 369–373 (1995).
34. Vogel-Sprott, M., Easdon, C., Fillmore, M., Finn, P., Justus, A.: Alcohol and behavioral control: cognitive and neural mechanisms. Alcohol Clin Exp Res 25, 117–121 (2001).
35. McGaugh, J. L.: The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annual review of neuroscience 27, 1–28, doi:10.1146/annurev.neuro.27.070203.144157 (2004).
36. Schuckit, M. A. et al.: A prospective evaluation of how a low level of response to alcohol predicts later heavy drinking and alcohol problems. The American journal of drug and alcohol abuse 37, 479–486, doi:10.3109/00952990.2011.598590 (2011).
37. Padula, C. B. et al.: Alcohol attenuates activation in the bilateral anterior insula during an emotional processing task: a pilot study. Alcohol and alcoholism 46, 547–552, doi:10.1093/alcalc/agr066 (2011).
38. Ray, S., Hanson, C., Hanson, S. J., Bates, M. E.: fMRI BOLD response in high-risk college students (Part 1): during exposure to alcohol, marijuana, polydrug and emotional picture cues. Alcohol and alcoholism 45, 437–443, doi:10.1093/alcalc/agq042 (2010).
39. Sripada, C. S., Angstadt, M., McNamara, P., King, A. C., Phan, K. L.: Effects of alcohol on brain responses to social signals of threat in humans. NeuroImage 55, 371–380, doi:10.1016/j.neuroimage.2010.11. 062 (2011).
40. Breiner, M. J., Stritzke, W. G., Lang, A. R.: Approaching avoidance. A step essential to the understanding of craving. Alcohol research & health : the journal of the National Institute on Alcohol Abuse and Alcoholism 23, 197–206 (1999).
41. Carter, C. S. et al.: Anterior cingulate cortex, error detection, and the online monitoring of performance. Science 280, 747–749 (1998).
42. Kril, J. J., Halliday, G. M., Svoboda, M. D., Cartwright, H.: The cerebral cortex is damaged in chronic alcoholics. Neuroscience 79, 983–998 (1997).
43. Anton, R. F., Moak, D. H., Latham, P.: The Obsessive Compulsive Drinking Scale: a self-rated instrument for the quantification of thoughts about alcohol and drinking behavior. Alcohol Clin Exp Res 19, 92–99 (1995).
44. Volkow, N. D. et al.: Effects of alcohol detoxification on dopamine D2 receptors in alcoholics: a preliminary study. Psychiatry Res 116, 163–172 (2002).
45. Gogh, V. v. Letter to Theo van Gogh. edited by Robert Harrison 513 (Written c. 22 July 1888 in Arles).
46. Agosti, V., Levin, F. R.: The effects of alcohol and drug dependence on the course of depression. Am J Addict 15, 71–75, doi:10.1080/ 10550490500419102 (2006).
47. Conway, K. P., Compton, W., Stinson, F. S., Grant, B. F.: Lifetime comorbidity of DSM-IV mood and anxiety disorders and specific drug use disorders: results from the National Epidemiologic Survey on Alcohol and Related Conditions. J Clin Psychiatry 67, 247–257 (2006).
48. Hillemacher T, Bleich S.: Alkoholabhängigkeit und komorbide psychische Störungen. In: Walter M, Gouzoulis-Mayfrank E (Hrsg.). Psychische Störungen und Suchterkrankungen. Diagnostik und Behandlung von Doppeldiagnosen. Stuttgart, Kohlhammer 2013, pp. 149–156.
&10 1/2014
PSYCHIATRIE NEUROLOGIE