

Transkript
FORTBILDUNG
Befunde funktionell bildgebender Studien bei Migräne
Bildgebende Studien haben unser Verständnis von Hirnfunktionsstörungen bei Migränepatienten erweitert und Erklärungsmodelle für die Entstehung von Migränesymptomen wie Kopfschmerz und Photophobie geliefert. Im Beitrag sind die Ergebnisse wichtig erscheinender Studien zusammengefasst, die das heutige Verständnis der Migränepathophysiologie stark beeinflusst haben.
Till Sprenger Michaela Andelova
von Till Sprenger, Michaela Andelova
D ie Migräne ist eine neurologische Erkrankung mit wiederholten Attacken von mittelstarken bis starken, typischerweise pulsierenden Kopfschmerzen, Licht-, Lärm- und Geruchsempfindlichkeit sowie Übelkeit (1). Die Prävalenz ist hoch, und die Erkrankung hat eine starke genetische Komponente. Initial ist der Verlauf der Migräne meist episodisch, das heisst, Kopfschmerzen treten an weniger als 15 Tagen im Monat auf. Im Verlauf kann jedoch eine Transformation mit einer Kopfwehhäufung im Sinne einer chronischen Migräne entstehen (d. h. 15 oder mehr Tage mit Kopfweh pro Monat). In den letzten zwei Dekaden wurde eine Vielzahl von Studien mit Positronenemissionstomografie (PET) und Magnetresonanztomografie (MRI) bei Migränepatienten durchgeführt. Diese Methoden erlauben Aussagen sowohl über die Hirnstruktur als auch die Funktion. In der Folge möchten wir einige uns wichtig erscheinende Studien zusammenfassen, welche unser heutiges Verständnis der Migränepathophysiologie beeinflusst haben.
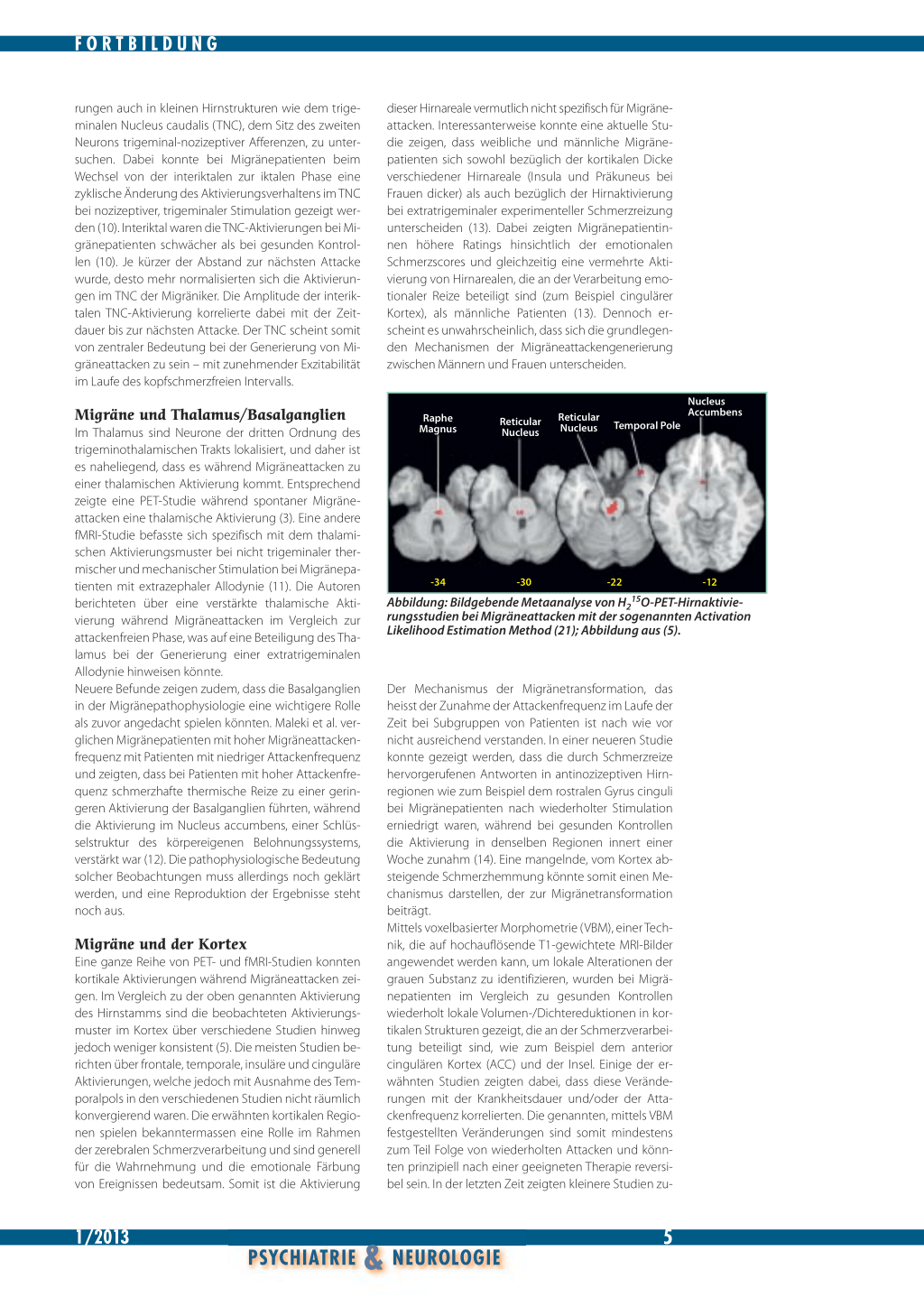
Hirnstamm- und Mittelhirnaktivierung bei Migräne Bereits in den Neunzigerjahren des letzten Jahrhunderts konnte eine wichtige PET-Studie einen erhöhten Blutfluss als Hinweis für eine neuronale Aktivierung im Hirnstamm und in weiteren supratentoriellen Hirnregionen während spontaner Migräneattacken nachweisen (2). Nach Therapie mit Sumatriptan war die Hirnstammaktivierung weiterhin nachweisbar, während die Aktivierungen in den Hemisphären nicht mehr vorhanden waren. Das wurde als Beleg für die kausale Bedeutung der Hirnstammaktivierung gewertet. Weitere PETStudien sowohl bei spontanen als auch bei experimentell herbeigeführten Migräneattacken führten bezüglich der genannten Hirnstammaktivierung zu ähnlichen Ergebnissen (Abbildung), (3, 4). Die Aktivierungen im Hirnstamm waren dabei überwiegend ipsilateral zur Kopfschmerzseite (4), was auf einen möglichen Zusammenhang zwischen Lateralisation der Schmerzen und lateralisierter Hirnstammdysfunktion hinwei-
sen könnte. Welche Hirnstammkerngebiete genau aktiviert werden, ist Gegenstand kontroverser Diskussionen. Die räumliche Auflösung der bisher in den Studien überwiegend verwendeten PET-Methodik lässt keine definitiven Aussagen zu (5). Verschiedene Autoren haben vermutet, dass die Aktivierungen unter anderem das periaquäduktale Höhlengrau (PAG), die Raphekerne und den Locus coeruleus betreffen (6). Diese Areale sind an der Schmerzverarbeitung sowie an der Modulation der kortikalen Exzitabilität beteiligt, und eine attackenartige Dysfunktion dieser Areale könnte möglicherweise eine episodenhafte Fehlverarbeitung sensorischer Reize erklären. Darüber hinaus wurden bei Migränepatienten in Hirnstammarealen auch strukturelle Veränderungen der grauen Substanz gezeigt (7). Auf der anderen Seite konnten neuere Studien mit funktioneller Magnetresonanztomografie zeigen, dass die genannten Hirnstammregionen während Migräneattacken sowohl durch nozizeptive als auch durch nicht nozizeptive Reize, zum Beispiel olfaktorische Stimuli, aktiviert werden. Die Hirnstammaktivierung scheint somit nicht spezifisch mit der Migräneschmerzverarbeitung zusammenzuhängen. In einer neueren PET-Aktivierungsstudie wurden sieben Migränepatienten im Gegensatz zu den oben genannten Untersuchungen bereits frühzeitig, das heisst innerhalb von vier Stunden nach Beginn spontaner Migräneattacken, untersucht. Die Autoren konnten eine Aktivierung nicht nur in der Brücke, sondern auch im Hypothalamus zeigen. Die Aktivierung des Hypothalamus persistierte ähnlich wie bei der ersten oben genannten Studie nach Kopfschmerzlinderung mit Sumatriptan (8). Eine ähnliche hypothalamische Aktivierung konnte kürzlich zudem noch früher, das heisst bereits während der Prodromalphase von Migräneattacken gezeigt werden (9). Eine hypothalamische Fehlfunktion könnte somit frühe Attackensymptome beziehungsweise Prodromalsymptome wie Heisshungergefühle, repetitives Gähnen, Müdigkeit und so weiter erklären. Die rasche Entwicklung von bildgebenden Verfahren, insbesondere im Rahmen der funktionellen Magnetresonanztomografie, ermöglicht es, neuerdings Aktivie-
&4 1/2013 PSYCHIATRIE NEUROLOGIE
FORTBILDUNG
rungen auch in kleinen Hirnstrukturen wie dem trigeminalen Nucleus caudalis (TNC), dem Sitz des zweiten Neurons trigeminal-nozizeptiver Afferenzen, zu untersuchen. Dabei konnte bei Migränepatienten beim Wechsel von der interiktalen zur iktalen Phase eine zyklische Änderung des Aktivierungsverhaltens im TNC bei nozizeptiver, trigeminaler Stimulation gezeigt werden (10). Interiktal waren die TNC-Aktivierungen bei Migränepatienten schwächer als bei gesunden Kontrollen (10). Je kürzer der Abstand zur nächsten Attacke wurde, desto mehr normalisierten sich die Aktivierungen im TNC der Migräniker. Die Amplitude der interiktalen TNC-Aktivierung korrelierte dabei mit der Zeitdauer bis zur nächsten Attacke. Der TNC scheint somit von zentraler Bedeutung bei der Generierung von Migräneattacken zu sein – mit zunehmender Exzitabilität im Laufe des kopfschmerzfreien Intervalls.
Migräne und Thalamus/Basalganglien Im Thalamus sind Neurone der dritten Ordnung des trigeminothalamischen Trakts lokalisiert, und daher ist es naheliegend, dass es während Migräneattacken zu einer thalamischen Aktivierung kommt. Entsprechend zeigte eine PET-Studie während spontaner Migräneattacken eine thalamische Aktivierung (3). Eine andere fMRI-Studie befasste sich spezifisch mit dem thalamischen Aktivierungsmuster bei nicht trigeminaler thermischer und mechanischer Stimulation bei Migränepatienten mit extrazephaler Allodynie (11). Die Autoren berichteten über eine verstärkte thalamische Aktivierung während Migräneattacken im Vergleich zur attackenfreien Phase, was auf eine Beteiligung des Thalamus bei der Generierung einer extratrigeminalen Allodynie hinweisen könnte. Neuere Befunde zeigen zudem, dass die Basalganglien in der Migränepathophysiologie eine wichtigere Rolle als zuvor angedacht spielen könnten. Maleki et al. verglichen Migränepatienten mit hoher Migräneattackenfrequenz mit Patienten mit niedriger Attackenfrequenz und zeigten, dass bei Patienten mit hoher Attackenfrequenz schmerzhafte thermische Reize zu einer geringeren Aktivierung der Basalganglien führten, während die Aktivierung im Nucleus accumbens, einer Schlüsselstruktur des körpereigenen Belohnungssystems, verstärkt war (12). Die pathophysiologische Bedeutung solcher Beobachtungen muss allerdings noch geklärt werden, und eine Reproduktion der Ergebnisse steht noch aus.
Migräne und der Kortex Eine ganze Reihe von PET- und fMRI-Studien konnten kortikale Aktivierungen während Migräneattacken zeigen. Im Vergleich zu der oben genannten Aktivierung des Hirnstamms sind die beobachteten Aktivierungsmuster im Kortex über verschiedene Studien hinweg jedoch weniger konsistent (5). Die meisten Studien berichten über frontale, temporale, insuläre und cinguläre Aktivierungen, welche jedoch mit Ausnahme des Temporalpols in den verschiedenen Studien nicht räumlich konvergierend waren. Die erwähnten kortikalen Regionen spielen bekanntermassen eine Rolle im Rahmen der zerebralen Schmerzverarbeitung und sind generell für die Wahrnehmung und die emotionale Färbung von Ereignissen bedeutsam. Somit ist die Aktivierung
dieser Hirnareale vermutlich nicht spezifisch für Migräneattacken. Interessanterweise konnte eine aktuelle Studie zeigen, dass weibliche und männliche Migränepatienten sich sowohl bezüglich der kortikalen Dicke verschiedener Hirnareale (Insula und Präkuneus bei Frauen dicker) als auch bezüglich der Hirnaktivierung bei extratrigeminaler experimenteller Schmerzreizung unterscheiden (13). Dabei zeigten Migränepatientinnen höhere Ratings hinsichtlich der emotionalen Schmerzscores und gleichzeitig eine vermehrte Aktivierung von Hirnarealen, die an der Verarbeitung emotionaler Reize beteiligt sind (zum Beispiel cingulärer Kortex), als männliche Patienten (13). Dennoch erscheint es unwahrscheinlich, dass sich die grundlegenden Mechanismen der Migräneattackengenerierung zwischen Männern und Frauen unterscheiden.
Raphe Magnus
Reticular Nucleus
Nucleus
Reticular
Accumbens
Nucleus Temporal Pole
-34 -30
-22
-12
Abbildung: Bildgebende Metaanalyse von H215O-PET-Hirnaktivierungsstudien bei Migräneattacken mit der sogenannten Activation Likelihood Estimation Method (21); Abbildung aus (5).
Der Mechanismus der Migränetransformation, das heisst der Zunahme der Attackenfrequenz im Laufe der Zeit bei Subgruppen von Patienten ist nach wie vor nicht ausreichend verstanden. In einer neueren Studie konnte gezeigt werden, dass die durch Schmerzreize hervorgerufenen Antworten in antinozizeptiven Hirnregionen wie zum Beispiel dem rostralen Gyrus cinguli bei Migränepatienten nach wiederholter Stimulation erniedrigt waren, während bei gesunden Kontrollen die Aktivierung in denselben Regionen innert einer Woche zunahm (14). Eine mangelnde, vom Kortex absteigende Schmerzhemmung könnte somit einen Mechanismus darstellen, der zur Migränetransformation beiträgt. Mittels voxelbasierter Morphometrie (VBM), einer Technik, die auf hochauflösende T1-gewichtete MRI-Bilder angewendet werden kann, um lokale Alterationen der grauen Substanz zu identifizieren, wurden bei Migränepatienten im Vergleich zu gesunden Kontrollen wiederholt lokale Volumen-/Dichtereduktionen in kortikalen Strukturen gezeigt, die an der Schmerzverarbeitung beteiligt sind, wie zum Beispiel dem anterior cingulären Kortex (ACC) und der Insel. Einige der erwähnten Studien zeigten dabei, dass diese Veränderungen mit der Krankheitsdauer und/oder der Attackenfrequenz korrelierten. Die genannten, mittels VBM festgestellten Veränderungen sind somit mindestens zum Teil Folge von wiederholten Attacken und könnten prinzipiell nach einer geeigneten Therapie reversibel sein. In der letzten Zeit zeigten kleinere Studien zu-
1/2013
&PSYCHIATRIE NEUROLOGIE
5
FORTBILDUNG
dem Veränderungen der kortikalen Dicke (Cortical Thickness) bei Migränepatienten. Konkret wurden eine Zunahme der kortikalen Dicke im primären somatosensorischen Kortex (15) sowie Veränderungen im visuellen Assoziationskortex (16) beobachtet. Demgegenüber konnte eine grössere Studie zur kortikalen Dicke diese Befunde nicht reproduzieren (17). Diese kontroversen Ergebnisse könnten auf Unterschiede der Studienpopulationen zurückzuführen sein, da insbesondere die Attackenfrequenz eine Rolle zu spielen scheint (18), welche sich zwischen den Studien deutlich unterschied.
Migräneaura und die Rolle des Kortex
Bezüglich der Migräneaura ist die pathophysiologische
Rolle des Kortex besser belegt als beim Migränekopf-
schmerz. Etwa ein Viertel der Migränepatienten berich-
ten über gelegentliche visuelle, somatosensorische,
aphasische oder motorische Auren, die typischerweise
dem Kopfschmerz vorausgehen. Bereits in den 1940er
Jahren hatten tierexperimentelle Beobachtungen von
Leao nahegelegt, dass die sogenannte Cortical Sprea-
ding Depression (CSD) (Reduktion der kortikalen
Aktivität, die sich mit langsamer Geschwindigkeit von
zirka 3 mm/min über den Kortex ausbreitet) das
elektrophysiologische Korrelat der visuellen Aura beim
Menschen darstellt. Es dauerte mehr als 50 Jahre bis
CSD-ähnliche Ereignisse mithilfe von elektrophysiolo-
gischen und bildgebenden Methoden beim Men-
schen nachgewiesen werden konnten. Am überzeu-
gendsten wurde dabei von Hadjikhani et al. mittels
fMRI bei einem Patienten mit provozierbarer Aura ge-
zeigt, dass visuell induzierte Signalfluktuationen mit
langsamer Frequenz über den okzipitalen Kortex wan-
dern, während sich gleichzeitig eine visuelle Aura in der
kontralateralen Gesichtsfeldhälfte ausbildete (19). Der
Ursprung dieser Welle von aura-assoziierten Signalalte-
rationen war im visuellen Assoziationskortex. Interes-
santerweise zeigte dieselbe Arbeitsgruppe später wie
oben genannt eine Zunahme der kortikalen Dicke bei
Migränepatienten im entsprechenden visuellen Asso-
ziationsareal (16). Aufgrund des Querschnitts-Studien-
designs war es dabei allerdings nicht möglich zu beur-
teilen, ob diese Veränderungen eher Ursachen oder
Folgen der Migräneaura sind. Eine Reproduktion des
genannten Befundes steht bis anhin aus. Neben der
genannten Abschwächung stimulusinduzierter Akti-
vierungen während der Aura wurde zudem mittels per-
fusionsgewichtetem MRI eine zerebrale Hypoperfusion
während Migräneauren im okzipitalen Kortex kontrala-
teral zu der betroffenen Seite gezeigt (20). Alles in allem
kann es als gesichert angesehen werden, dass CSD-
ähnliche Ereignisse die pathophysiologische Grund-
lage von Migräneauren darstellen.
●
Korrespondenzadresse:
Prof. Dr. med. Till Sprenger
Universitätsspital Basel
Neurologische Klinik und Poliklinik
Petersgraben 4
4031 Basel
Tel. 061-328 63 86
Fax 061-265 41 62
E-Mail: till.sprenger@usb.ch
Merksätze:
● Während Migräneattacken konnte konsis-
tent eine Aktivierung von Hirnstammzen-
tren gezeigt werden. Betroffen sind vermut-
lich antinozizeptive Kerngebiete wie das pe-
riaquäduktale Grau.
● Die bei Migräne gelegentlich beobachtete
extratrigeminale Allodynie entsteht ver-
mutlich durch thalamische Sensitivierung.
● Auch kortikale Areale werden während Mi-
gräneattacken aktiviert, allerdings ist das
beobachtete Aktivierungsmuster über ver-
schiedene Studien hinweg bis anhin nicht
konsistent.
● Bildgebend konnte analog zur elektro-
physiologisch im Tiermodell nachweisbaren
Cortical Spreading Depression belegt wer-
den, dass es während der Migräneaura zu ei-
ner sich langsam über den Okzipitalkortex
ausbreitenden Welle mit Depression der
kortikalen Erregbarkeit kommt.
Referenzen: 1. The International Classification of Headache Disorders: 2nd edition.
Cephalalgia. 2004; 24 Suppl 1: 9–160. 2. Weiller C, May A, Limmroth V, Juptner M, Kaube H, Schayck RV, et al.:
Brain stem activation in spontaneous human migraine attacks. Nat Med. 1995; 1(7): 658–60. 3. Afridi SK, Giffin NJ, Kaube H, Friston KJ, Ward NS, Frackowiak RS, et al.: A positron emission tomographic study in spontaneous migraine. Arch Neurol. 2005; 62(8): 1270–5. 4. Afridi SK, Matharu MS, Lee L, Kaube H, Friston KJ, Frackowiak RS, et al.: A PET study exploring the laterality of brainstem activation in migraine using glyceryl trinitrate. Brain. 2005; 128(Pt 4): 932–9. 5. Sprenger T, Borsook D.: Migraine changes the brain: neuroimaging makes its mark. Curr Opin Neurol. 2012; 25(3): 252–62. 6. Sprenger T, Goadsby PJ.: Migraine pathogenesis and state of pharmacological treatment options. BMC Med. 2009; 7: 71. 7. Rocca MA, Ceccarelli A, Falini A, Colombo B, Tortorella P, Bernasconi L, et al.: Brain gray matter changes in migraine patients with T2-visible lesions: a 3-T MRI study. Stroke: 2006; 37(7): 1765–70. 8. Denuelle M, Fabre N, Payoux P, Chollet F, Geraud G.: Hypothalamic activation in spontaneous migraine attacks. Headache. 2007; 47(10): 1418–26. 9. Sprenger T, Maniyar F, Monteith T, Schankin C, Goadsby P.: Midbrain Activation in the Premonitory Phase of Migraine: A H215O-PET study. Headache. 2012; 52: 863. 10. Stankewitz A, Aderjan D, Eippert F, May A.: Trigeminal nociceptive transmission in migraineurs predicts migraine attacks. The Journal of neuroscience: 2011; 31(6): 1937–43. 11. Burstein R, Jakubowski M, Garcia-Nicas E, Kainz V, Bajwa Z, Hargreaves R, et al.: Thalamic sensitization transforms localized pain into widespread allodynia. Annals of neurology. 2010; 68(1): 81–91. 12. Maleki N, Becerra L, Nutile L, Pendse G, Brawn J, Bigal M, et al.: Migraine attacks the Basal Ganglia. Mol Pain. 2011; 7: 71. 13. Maleki N, Linnman C, Brawn J, Burstein R, Becerra L, Borsook D.: Her versus his migraine: multiple sex differences in brain function and structure. Brain: 2012; 135(Pt 8): 2546–59. 14. Aderjan D, Stankewitz A, May A.: Neuronal mechanisms during repetitive trigemino-nociceptive stimulation in migraine patients. Pain. 2010; 151(1): 97–103. 15. DaSilva AF, Granziera C, Snyder J, Hadjikhani N.: Thickening in the somatosensory cortex of patients with migraine. Neurology. 2007; 69(21): 1990–5. 16. Granziera C, DaSilva AF, Snyder J, Tuch DS, Hadjikhani N.: Anatomical alterations of the visual motion processing network in migraine with and without aura. PLoS Med. 2006; 3(10): e402. 17. Datta R, Detre JA, Aguirre GK, Cucchiara B.: Absence of changes in cortical thickness in patients with migraine. Cephalalgia: 2011; 31(14): 1452–8. 18. Maleki N, Becerra L, Brawn J, Bigal M, Burstein R, Borsook D.: Concurrent functional and structural cortical alterations in migraine. Cephalalgia: 2012; 32(8): 607–20. 19. Hadjikhani N, Sanchez Del Rio M, Wu O, Schwartz D, Bakker D, Fischl B, et al.: Mechanisms of migraine aura revealed by functional MRI in human visual cortex. Proc Natl Acad Sci USA. 2001; 98(8): 4687–92. 20. Sanchez del Rio M, Bakker D, Wu O, Agosti R, Mitsikostas DD, Ostergaard L, et al.: Perfusion weighted imaging during migraine: spontaneous visual aura and headache. Cephalalgia. 1999; 19(8): 701–7. 21. Eickhoff SB, Laird AR, Grefkes C, Wang LE, Zilles K, Fox PT.: Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: a random-effects approach based on empirical estimates of spatial uncertainty. Hum Brain Mapp. 2009; 30(9): 2907–26.
&6 1/2013 PSYCHIATRIE NEUROLOGIE