



Transkript
NUTRIGENOMIK
Nutri-Epigenetik: Der Zusammenhang zwischen Ernährung und Genetik
Doreen Gille und Guy Vergères
Die Art und Weise, wie wir uns ernähren, kann die Gesundheit unserer Kinder und Enkelkinder beeinflussen. Denn es gibt immer mehr Hinweise dafür, dass unsere Ernährung in epigenetische Mechanismen eingreifen kann. Im Beitrag wird der Zusammenhang zwischen Ernährungsmustern und der folgenden Beeinflussung epigenetischer Modifikationen dargestellt.
Doreen Gille
Als im Jahr 2001 Wissenschaftler verkündeten (1), das Geheimnis der Menschheit – nämlich die Entschlüsselung des Humangenoms – sei gelüftet, war die Aufregung gross. Nur kurze Zeit später wurde jedoch deutlich, dass das Wissen über die Sequenz des menschlichen Genoms nicht ausreicht, um zu verstehen, wie verschiedenste Prozesse im Körper funktionieren. Auch wenn die individuelle genetische Information bis auf wenige zufällige Veränderungen (Mutationen) ein Leben lang gleich bleibt, gibt es Modifikationen, strukturelle Umwandlungen und chemische Reaktionen, die unsere Genaktivität so beeinflussen, dass man als Individuum nicht mehr genau so ist wie durch die genetische Information definiert. Beispielsweise können bestimmte Bereiche des Erbguts (Gene) stillgelegt, andere wiederum aktiviert oder für diverse wichtige Enzyme leichter zugänglich gemacht werden. Diese Veränderungen, die als epigenetische Modifikationen bezeichnet werden, haben Einfluss darauf, wie unser Körper bis ins kleinste Detail funktioniert und reagiert. Arthur Riggs, ein Vorreiter in der Erforschung epigenetischer Veränderungen, hat dieses Phänomen 1996 treffend als die «vererbbaren Veränderungen der Genaktivität ohne Veränderung der Abfolge der DNABausteine» formuliert. Epigenetische Modifikationen sind reversibel, können aber bei der Teilung von Körperzellen weitervererbt werden. Funktionell spielen sie zum Beispiel eine Rolle bei der Entwicklung des Organismus, der Spezialisierung verschiedener Zelltypen und bei der Entstehung beziehungsweise Prävention verschiedener Krankheiten (2). Vereinfacht beschrieben beschäftigt sich die Epigenetik mit Mechanismen der Genregulation, indem sie die Verpackung der DNA (Desoxyribonukleinsäure) in der Zelle genauer betrachtet; die DNA ist in jeder Körperzelle gleich, aber ihre Verpackung nicht. Generell unterscheidet man drei epigenetische Mechanismen:
1. chemische Veränderungen der DNA; 2. chemische Veränderungen, der an die DNA ge-
bundenen Proteine (Histone) und 3. den Einfluss von RNA-Molekülen.
Chemische Veränderungen der DNA (DNA-Methylierungen)
Um den Ansatz der DNA-Methylierungen verstehen
zu können, muss nachvollziehbar sein, wie genau die
DNA in der Zelle organisiert ist (Abbildung 1). Von
einer DNA-Methylierung spricht man, wenn sich
kleine Moleküle (sogenannte Methylgruppen: -CH3)
Guy Vergères
kovalent an die DNA anheften, wodurch die Chroma-
tinstruktur verändert und folglich die Aktivität von
Genen beeinflusst wird. Vereinfacht gesagt wird ein
definiertes Gen durch diese Methylierung stillgelegt
und die enthaltene Information zur Herstellung eines
Proteins verschlossen, sodass das Protein nicht mehr
hergestellt werden kann. Dieses Phänomen bezeich-
net man als «gene silencing». Die Konse-
quenzen solcher Modifikationen sind komplex, und je nach Position der Methylierung im Erbgut, kann Genexpression auch durch Methylierung aktiviert wer-
Nutri-épigénétique: la relation entre alimentation et génétique
den. DNA-Methylierungen sind also keineswegs nachteilig, sondern vielmehr essenziell für eine normale Entwicklung der Körperzellen. Denn eine Deregulierung der DNA-Methylierung kann schwerwie-
Mots clés: modifications des histones – régulation épigénétique – modèle alimentaire et épigénétique
gende Krankheiten wie zum Beispiel Krebs, Diabetes, neurologische Erkrankungen und so weiter zur Folge haben. Häufig gehen diese meist chronischen und altersbedingten Leiden mit Hypomethylierungen einher, die durch eine zu geringe Aufnahme von methylgruppenhaltigen Lebensmitteln (siehe Absatz Nutri-Epigenetik) oder eine altersbedingte, reduzierte
De plus en plus d’indices suggèrent que notre alimentation peut intervenir sur des mécanismes épigénétiques. La présentation de Doreen Gille et Guy Vergères, Institut des sciences alimentaires, présente les relations entre des modèles alimentaires et les effets sur les modifications épigénétiques qui en résultent.
3|2016 SZE 9
NUTRIGENOMIK
© fotolia.com – designua
Chemische Veränderungen der mit der DNA verbundenen Proteine (Histonmodifikationen)
Auch im Fall der Histone (Kasten 1) können sich verschiedene Moleküle, wie zum Beispiel Methylgruppen, anlagern. Sie sorgen dafür, dass die Struktur des Chromatins wesentlich kompakter ist und somit die Möglichkeit, die Information der Gene abzuschreiben, enorm erschwert wird («gene silencing»). Die Anlagerung von Acetyl- und Phosphorgruppen hingegen öffnet die Chromatinstruktur und erhöht die Zugänglichkeit zur DNA, was in der Folge die Genexpression aktiviert (6). Die Auswirkungen dieser Modifikationen auf die Genexpression sind ebenfalls komplex, und die Forschung ist stetig damit konfrontiert, dieses vereinfachte Modell mit der experimentellen Realität in Einklang zu bringen.
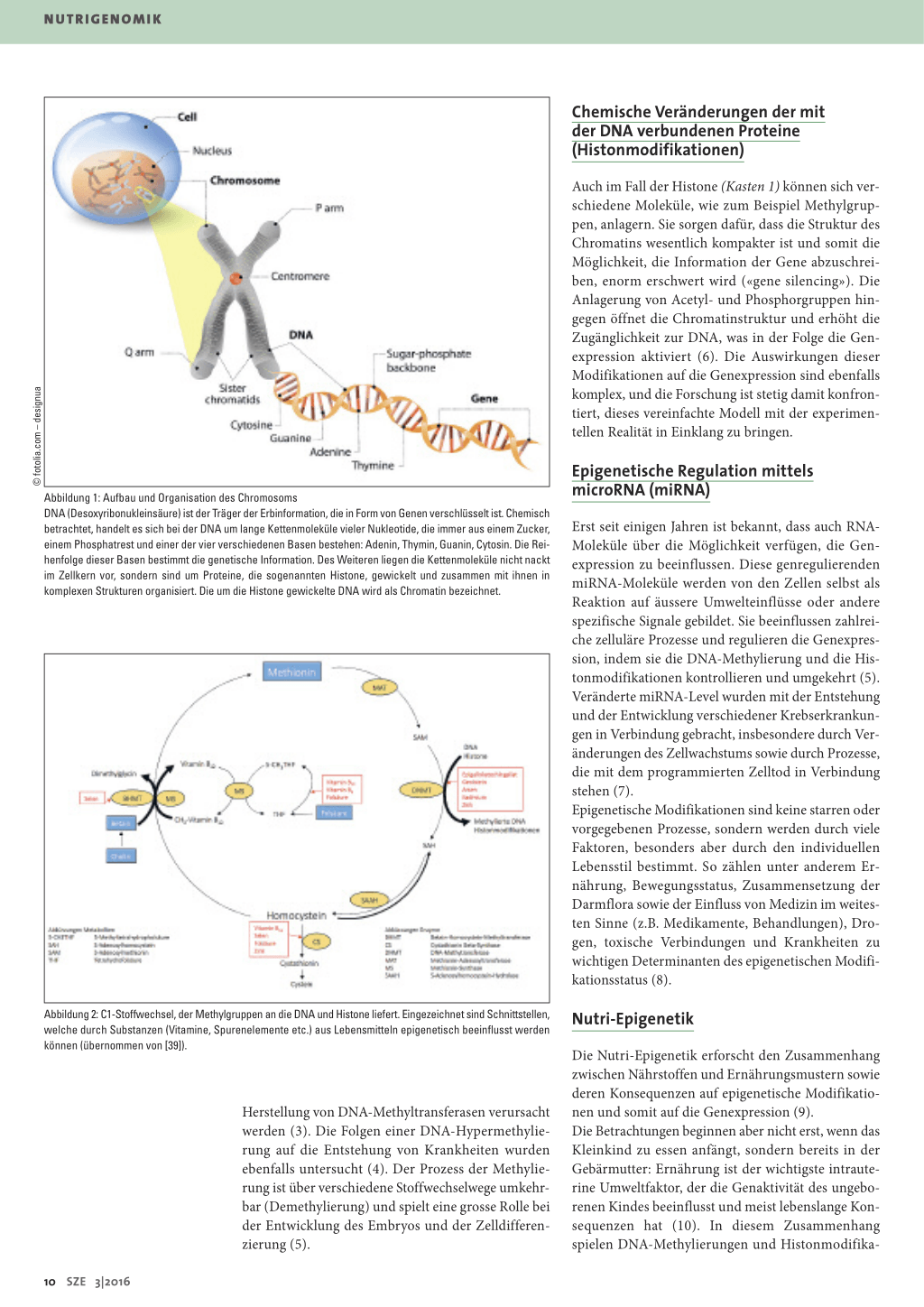
Abbildung 1: Aufbau und Organisation des Chromosoms DNA (Desoxyribonukleinsäure) ist der Träger der Erbinformation, die in Form von Genen verschlüsselt ist. Chemisch betrachtet, handelt es sich bei der DNA um lange Kettenmoleküle vieler Nukleotide, die immer aus einem Zucker, einem Phosphatrest und einer der vier verschiedenen Basen bestehen: Adenin, Thymin, Guanin, Cytosin. Die Reihenfolge dieser Basen bestimmt die genetische Information. Des Weiteren liegen die Kettenmoleküle nicht nackt im Zellkern vor, sondern sind um Proteine, die sogenannten Histone, gewickelt und zusammen mit ihnen in komplexen Strukturen organisiert. Die um die Histone gewickelte DNA wird als Chromatin bezeichnet.
Epigenetische Regulation mittels microRNA (miRNA)
Erst seit einigen Jahren ist bekannt, dass auch RNAMoleküle über die Möglichkeit verfügen, die Genexpression zu beeinflussen. Diese genregulierenden miRNA-Moleküle werden von den Zellen selbst als Reaktion auf äussere Umwelteinflüsse oder andere spezifische Signale gebildet. Sie beeinflussen zahlreiche zelluläre Prozesse und regulieren die Genexpression, indem sie die DNA-Methylierung und die Histonmodifikationen kontrollieren und umgekehrt (5). Veränderte miRNA-Level wurden mit der Entstehung und der Entwicklung verschiedener Krebserkrankungen in Verbindung gebracht, insbesondere durch Veränderungen des Zellwachstums sowie durch Prozesse, die mit dem programmierten Zelltod in Verbindung stehen (7). Epigenetische Modifikationen sind keine starren oder vorgegebenen Prozesse, sondern werden durch viele Faktoren, besonders aber durch den individuellen Lebensstil bestimmt. So zählen unter anderem Ernährung, Bewegungsstatus, Zusammensetzung der Darmflora sowie der Einfluss von Medizin im weitesten Sinne (z.B. Medikamente, Behandlungen), Drogen, toxische Verbindungen und Krankheiten zu wichtigen Determinanten des epigenetischen Modifikationsstatus (8).
Abbildung 2: C1-Stoffwechsel, der Methylgruppen an die DNA und Histone liefert. Eingezeichnet sind Schnittstellen, welche durch Substanzen (Vitamine, Spurenelemente etc.) aus Lebensmitteln epigenetisch beeinflusst werden können (übernommen von [39]).
Herstellung von DNA-Methyltransferasen verursacht werden (3). Die Folgen einer DNA-Hypermethylierung auf die Entstehung von Krankheiten wurden ebenfalls untersucht (4). Der Prozess der Methylierung ist über verschiedene Stoffwechselwege umkehrbar (Demethylierung) und spielt eine grosse Rolle bei der Entwicklung des Embryos und der Zelldifferenzierung (5).
Nutri-Epigenetik
Die Nutri-Epigenetik erforscht den Zusammenhang zwischen Nährstoffen und Ernährungsmustern sowie deren Konsequenzen auf epigenetische Modifikationen und somit auf die Genexpression (9). Die Betrachtungen beginnen aber nicht erst, wenn das Kleinkind zu essen anfängt, sondern bereits in der Gebärmutter: Ernährung ist der wichtigste intrauterine Umweltfaktor, der die Genaktivität des ungeborenen Kindes beeinflusst und meist lebenslange Konsequenzen hat (10). In diesem Zusammenhang spielen DNA-Methylierungen und Histonmodifika-
10 SZE 3|2016
NUTRIGENOMIK
tionen eine wichtige Rolle. Die Ernährung der Mutter und dadurch auch die Ernährung des Fötus hat viele entwicklungsspezifische Anpassungen beim Nachwuchs zur Folge, besonders auf der Ebene des Stoffwechsels. Damit werden bereits die Weichen für mögliche Krankheiten kardiovaskulären, metabolischen oder endokrinen Ursprungs im Erwachsenenalter gestellt. Eine optimale Ernährung der Mutter sichert die optimale Entwicklung des Fötus, vor allem aber reduziert sich damit auch das Risiko, im Alter an chronischen Krankheiten zu leiden (11, 12). Die Kombination dieser Faktoren bedingt also zu einem grossen Teil, ob wir ein erhöhtes Risiko haben, bestimmte Erkrankungen im Erwachsenenalter zu entwickeln. Denn all diese Faktoren modifizieren unsere epigenetischen Merkmale (5). Wie schon erwähnt, sind epigenetische Veränderungen nicht starr, sondern empfänglich für Veränderungen, die zum Beispiel von der Diät und dem individuellen Lebensstil hervorgerufen werden können. Krebshemmende Eigenschaften werden unter anderem verschiedenen bioaktiven Lebensmittelkomponenten zugeschrieben, die wiederum Methylierungsmuster und andere epigenetische Veränderungen regulieren können wie: 1. die Aktivität des Enzyms DNA-Methyltransferase
(überträgt Methylgruppen auf die DNA) und Histonacetyltransferase (überträgt Acetylgruppen auf Histone) 2. DNA-Demethylierungsaktivität (Entfernung von Methylgruppen) und Histondeacetylase (Entfernung von Acetylgruppen) 3. Transkriptionsfaktoren, die das Abschreiben der Gene aktivieren oder deaktivieren 4. chromatinverändernde Eigenschaften (5).
Nährstoffe, die die DNA-Methylierung und die Histonmodifikationen beeinflussen
Die Interaktion zwischen Nährstoffen und DNA- beziehungsweise Histonmethylierungen beruht auf einem zentralen Prozess im Körper des Menschen, dem C1-Stoffwechsel (Abbildung 2), in dessen Zentrum Folsäure steht und der grundsätzlich für die Übertragung von Kohlenstoffatomen und somit auch Methylgruppen (-CH3) wichtig ist. Zwei Verbindungen des C1-Stoffwechsels beeinflussen die Methylierung von DNA und Histonen besonders: S-Adenosylmethionin (spendet Methylgruppen) und S-Adenosylhomocystein (hemmt Enzyme, die Methylgruppen übertragen). Jeder Nahrungsbestandteil, der auf diese beiden Schlüsselstellen einwirken kann, stellt einen wichtigen Einflussfaktor für Methylierungsreaktionen dar. Verschiedene Tierstudien, aber auch Untersuchungen im Menschen zeigen, dass folsäurereiche Lebensmittel einen protektiven Effekt bezüglich der Entwicklung von Darmkrebs ausüben. Dies wahrscheinlich aufgrund ihrer Rolle bei der DNA-Synthese und -Methylierung. Es gibt allerdings auch Hinweise, dass besonders hohe Konzentrationen an Folsäure die Ent-
stehung von Tumoren begünstigen. Noch ist nicht abschliessend geklärt, was bei einem Folsäuremangel im Körper passiert. Es wird aber davon ausgegangen, dass der Mangel zu Hypomethylierungen der DNA führt und die Histonmethylierung beeinflusst werden könnte (13). Folsäurehaltige Lebensmittel sind vor allem Endivie, Spargel, Kohl, Spinat, Vollkornprodukte und Weizenkleie, Erd- und Walnüsse, Eigelb und Leber (14). Weitere wesentliche Faktoren in diesem (C1-)Prozess sind die Nährstoffe Vitamin B12, Betain, Methionin und Cholin. Denn diese Nährstoffe tragen ebenfalls Methylgruppen (5). Daher geht man davon aus, dass eine erhöhte Aufnahme dieser Verbindungen DNAund Histonmethylierungen gewährleisten könnte. Reich an methylgruppenhaltigen Substanzen sind vor allem tierische Produkte wie Milchprodukte, Fleisch und Fleischprodukte sowie Ei, aber auch Getreideerzeugnisse, Kartoffeln, Linsen, Bohnen, Erbsen, Nüsse, Sojamehl, Bierhefe, Rübenpflanzen und Weizenkleie/Weizenkeime (15–18).
Ernährungsmuster beeinflussen epigenetische Modifikationen
Auch für andere bioaktive Komponenten in Lebensmitteln konnte vor allem in Tierstudien Potenzial für epigenetische Veränderungen identifiziert werden. Wasserlösliche B-Vitamine wie zum Beispiel Biotin, Niacin und Pantothensäure spielen eine wichtige Rolle bei Histonmodifikationen, indem sie die Aktivität verschiedener Enzyme beeinflussen (19). Der Bereich der Histonacetylierung wurde bereits breit beforscht: Auch Verbindungen wie Resveratrol (Rotwein), Buttersäure (Stoffwechselprodukt vom Ballaststoffverdau durch die Mikroflora), Sulforaphan (Kohl, Broccoli) und Diallylsulfid (Knoblauch) inhibieren das Enzym Histondeactelyase (20–22), während Curcurmin (Gelbwurzel) die Histonacetyltransferase hemmt (23) – beides gewünschte Effekte und mögliche therapeutische Potenziale in der Krebstherapie (5). In Bezug auf Interaktionen zwischen Nährstoffen beziehungsweise bioaktiven Lebensmittelkomponenten und Histonmethylierungen sind kaum Informationen bekannt, obwohl diese Veränderungen stark mit dem Alterungsprozess und Krebserkrankungen einhergehen (24). Genistein (Sojabohne) und Catechin (Tee) beeinträchtigen das Enzym DNA-Methyltransferase, wodurch es zu einer Hypomethylierung von Genen kommt, die mit Krebs in Verbindung gebracht werden (25–27). Allerdings muss auch erwähnt werden, dass Hypomethylierungen nur für bestimmte krebsassoziierte Gene günstig sind, während das Risiko, an anderen Krebsarten zu erkranken, durch eine solche Hypomethylierung erhöht sein kann (28). Ausserdem sollte beachtet werden, dass die meisten dieser Erkenntnisse aus In-vitro- oder Tierstudien stammen und die Relevanz für den Menschen noch genauer untersucht werden muss. Nun konsumiert der Mensch nicht nur Nährstoffe,
Kasten 1:
So wird aus Geninformation ein Protein
Wird ein «Gen exprimiert», dann wird dieser Genabschnitt auf der DNA geöffnet, und es kommt zu einem «Ableseprozess». Dabei entsteht mit der DNA als Vorbild ein neues Molekül, die RNA (Ribonukleinsäure). Die RNA ist beweglich, verlässt den Zellkern, und entsprechend ihrer Information (abgeschrieben von der DNA) wird dann das passende Protein gebildet. Diese Prozesse bezeichnet man als Transkription (abschreiben) und Translation (übersetzen) und gesamthaft als Genexpression.
Kasten 2:
Kurz & knapp: Epigenetik, Epigenomik und Nutri-Epigenetik
Epigenetik (epi = griechisch für «dazu»/«ausserdem», genetik = griechisch für «Abstammung»/ «Ursprung») beschäftigt sich mit Veränderungen der DNA-Sequenz, die nicht auf Mutationen beruhen und dennoch weitergegeben werden. Die Veränderungen lassen sich im Phänotyp, aber nicht im Genotyp beobachten. Die Disziplin Epigenetik beschäftigt sich mit Modifikationen bei einzelnen Genen oder Gensets, während die Epigenomik epigenetische Akteure im Gesamtgenom untersucht. NutriEpigenetik erforscht den Einfluss von Nährstoffen und Ernährungsmustern auf epigenetische Veränderungen.
3|2016 SZE 11
NUTRIGENOMIK
«Epigenetische
Modifikationen sind reversibel, können aber bei der Teilung von Körperzellen wei-
»tervererbt werden.
sondern Lebensmittel entsprechend seinem persönlichen Ernährungsmuster. Die allgemeine Ernährungsweise kann den epigenetischen Status eines Genoms beeinflussen, sodass eine grosse Anzahl an Genen in metabolischen und physiologischen Prozessen und Systemen umprogrammiert wird. Das hat einen Einfluss auf typische Erkrankungen, die mit dem Alterungsprozess einhergehen können, wie Diabetes Typ 2, koronare Herzkrankheiten, neurodegenerative Erkrankungen und Krebs. So konnte zum Beispiel bei trächtigen Mäusen gezeigt werden, dass proteinarme Diäten Bluthochdruck, Fettstoffwechselstörungen und einen gestörten Glukosestoffwechsel in den Nachkommen provozieren, da die DNA-Methylierung und die Histonmodifikationen spezifische Schlüsselgene wie den Glukokortikoidrezeptor und die PPARa (Peroxisomen-Proliferator-aktivierte Rezeptoren) betrafen (29, 30). Auch die Bildung von miRNA ist zu einem grossen Teil vom Methylierungsstatus der DNA sowie von der Ausprägung der Histonmodifikation abhängig. In einem Experiment wurde gezeigt, dass eine methylgruppenarme Diät eine veränderte Expression verschiedener miRNA zur Folge hatte und die Versuchstiere schliesslich an Krebs erkrankten oder eine nicht alkoholbedingte Fettleber entwickelten (31, 32). Eine ähnliche Beobachtung wurde bei einem Folsäuremangel des Menschen gemacht (33). Ausserdem konnte gezeigt werden, dass eine cholesterinreiche Diät im Vergleich zu einer Standarddiät das Körpergewicht, das Gesamtcholesterol und das HDL in adipösen Minipigs ansteigen liess, während die Expression einer speziellen miRNA (miRNA122) reduziert war (34). Das bedeutet, dass diese miRNA im Zusammenhang mit Adipositas steht. Bis anhin wurden verschiedene bioaktive Lebensmittelkomponenten identifiziert, die mit einer Regulierung der miRNA einhergehen und damit das Risiko für die Entstehung von Krebs reduzieren: Genistein (35), Curcumin (36) und Retinol (in tierischen Lebensmitteln und als Carotinoid in gelborangem Gemüse) (37).
Schlussfolgerung
Während des gesamten Lebens können Mikro- und Makronährstoffe physiologische und pathophysiologische Prozesse über epigenetische Modifikationen und über die Genexpression steuern und beeinflussen. Diese Erkenntnis und die damit einhergehenden Forschungsergebnisse offenbaren einmal mehr die Wirkung einer ausgewogenen und abwechselungsreichen Ernährungsweise auf den Erhalt der Gesundheit beziehungsweise auf die Vorbeugung von Krankheiten. Dabei ist es nicht möglich, Effekte nur bestimmten Nährstoffen oder bioaktiven Lebensmittelkomponenten zuzuordnen, weil diese auch direkt mit Genen und anderen Nährstoffen interagieren können. Die Komplexität dieser Wechselwirkungen wird nochmals erhöht, indem sich auch die epigenetischen Mechanismen gegenseitig beeinflussen.
Zudem können auch epigenetische Modifikationen, die in Keimzellen auftreten, auf folgende Generationen transferiert werden (38). Dieses Phänomen zeigt die erhebliche Konsequenz des Lebensstils der Elterngeneration auf ihre Nachkommen (9). Denn die Art und Weise, wie wir uns ernähren, kann die Gesundheit unserer Kinder und Enkelkinder beeinflussen! Doch speziell auf diesem Gebiet benötigt es sowohl weitere mechanistische Grundlagen via Tiermodell als auch experimentelle Evidenz beim Menschen, um den reellen transgenerationalen Einfluss der Ernährung auf die Humangesundheit genauer beurteilen zu können. Weitere Forschung auf diesem Gebiet ist daher unerlässlich.
Korrespondenzadresse: Doreen Gille Agroscope Institut für Lebensmittelwissenschaften Schwarzenburgstrasse 161 3003 Bern E-Mail: doreen.gille@agroscope.admin.ch Internet: www.agroscope.ch
Literatur: 1. Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J et al.: Initial sequencing and analysis of the human genome. Nature. 2001; 409: 860–921. 2. Fraga MF, Ballestar E, Villar-Garea A, Boix-Chornet M, Espada J, Schotta et al.: Loss of acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a common hallmark of human cancer. Nature genetics. 2005; 37: 391–400. 3.Li Y, Liu Y, Strickland FM, Richardson B: Age-dependent decreases in DNA methyltransferase levels and low transmethylation micronutrient levels synergize to promote overexpression of genes implicated in autoimmunity and acute coronary syndromes. Experimental gerontology. 2010; 45: 312–22. 4. Rodenhiser D, Mann M: Epigenetics and human disease: translating basic biology into clinical applications. CMAJ: Canadian Medical Association journal (journal de l’association medicale canadienne). 2006; 174: 341–348. 5. Choi SW, Friso S: Epigenetics: A new bridge between nutrition and health. Advances in nutrition. 2010; 1: 8–16. 6. Jenuwein T, Allis CD: Translating the histone code. Science. 2001; 293: 1074–1080. 7. Iorio MV, Piovan C, Croce CM: Interplay between microRNAs and the epigenetic machinery: an intricate network. Biochimica et biophysica acta. 2010; 1799: 694–701. 8. Feil R, Fraga MF: Epigenetics and the environment: emerging patterns and implications. Nature reviews genetics. 2011; 13: 97–109. 9. Vergères G, Gille D: Nutri(epi)genomik. In: Lux V, Richter T, editors. Kulturen der Epigenetik: vererbt, codiert, übertragen. Berlin: de Gruyter; 2014. p.1–9. 10. Chen JH, Tarry-Adkins JL, Matharu K, Yeo GS, Ozanne SE: Maternal protein restriction affects gene expression profiles in the kidney at weaning with implications for the regulation of renal function and lifespan. Clinical science. 2010; 119: 373–84. 11. Geraghty AA, Lindsay KL, Alberdi G, McAuliffe FM, Gibney ER: Nutrition during pregnancy impacts offspring’s epigenetic status – evidence from human and animal studies. Nutrition and metabolic insights. 2015; 8: 41–47. 12. Sinclair KD, Allegrucci C, Singh R, Gardner DS, Sebastian S, Bispham J et al.: DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104: 19351–19356. 13. Garcia BA, Luka Z, Loukachevitch LV, Bhanu NV, Wagner C: Folate deficiency affects histone methylation. Medical hypotheses. 2016; 88: 63–67. 14. Druml W, Stein J: Mikronährstoffe. In: Stein J, Jauch KW, editors. Praxishandbuch klinische Ernährung und Infusionstherapie. Berlin Heidelberg: Springer-Verlag; 2013. p. 97. 15. Burgerstein UP, Schurgast H, Zimmermann MB: Handbuch Nährstoffe: Vorbeugen und heilen durch ausgewogene Ernährung: TRIAS; 2012.
12 SZE 3|2016
16. Gille D, Schmid A: Vitamin B12 in meat and dairy products. Nutrition reviews. 2015; 73: 106–115. 17. Zeisel SH, Mar MH, Howe JC, Holden JM: Concentrations of cholinecontaining compounds and betaine in common foods. Journal of nutrition. 2003; 133: 1302–1307. 18. Druml W, Stein J: Ernährung bei Krankheiten der Niere. In: Stein J, Jauch KW, editors. Praxishandbuch klinische Ernährung und Infusionstherapie. Berlin Heidelberg: Springer-Verlag; 2013. p. 536. 19. Kirkland JB: Niacin status impacts chromatin structure. Journal of nutrition. 2009; 139: 2397–2401. 20. Do DP, Pai SB, Rizvi SA, D’Souza MJ: Development of sulforaphaneencapsulated microspheres for cancer epigenetic therapy. International journal of pharmaceutics. 2010; 386: 114–121. 21. Druesne-Pecollo N, Chaumontet C, Pagniez A, Vaugelade P, Bruneau A, Thomas M et al.: In vivo treatment by diallyl disulfide increases histone acetylation in rat colonocytes. Biochemical and biophysical research communications. 2007; 354: 140–147. 22. Kuroiwa-Trzmielina J, de Conti A, Scolastici C, Pereira D, Horst MA, Purgatto E et al.: Chemoprevention of rat hepatocarcinogenesis with histone deacetylase inhibitors: efficacy of tributyrin, a butyric acid prodrug. International journal of cancer. 2009; 124: 2520–2527. 23. Marcu MG, Jung YJ, Lee S, Chung EJ, Lee MJ, Trepel J, Neckers L: Curcumin is an inhibitor of p300 histone acetylatransferase. Medicinal chemistry. 2006; 2: 169–174. 24. Pogribny IP, Ross SA, Tryndyak VP, Pogribna M, Poirier LA, Karpinets TV: Histone H3 lysine 9 and H4 lysine 20 trimethylation and the expression of Suv4-20h2 and Suv-39h1 histone methyltransferases in hepatocarcinogenesis induced by methyl deficiency in rats. Carcinogenesis. 2006; 27: 1180–1186. 25. Fang M, Chen D, Yang CS: Dietary polyphenols may affect DNA methylation. Journal of nutrition. 2007; 137: 223s–228s. 26. Qin W, Zhu W, Shi H, Hewett JE, Ruhlen RL, MacDonald RS et al.: Soy isoflavones have an antiestrogenic effect and alter mammary promoter hypermethylation in healthy premenopausal women. Nutrition and cancer. 2009; 61: 238–244. 27. Raynal NJ, Charbonneau M, Momparler LF, Momparler RL: Synergistic effect of 5-Aza-2’-deoxycytidine and genistein in combination against leukemia. Oncology research. 2008; 17: 223–230. 28. Tang WY, Newbold R, Mardilovich K, Jefferson W, Cheng RY, Medvedovic M, Ho SM: Persistent hypomethylation in the promoter of nucleosomal binding protein 1 (Nsbp1) correlates with overexpression of Nsbp1 in mouse uteri neonatally exposed to diethylstilbestrol or genistein. Endocrinology. 2008; 149: 5922–5931.
29. Lillycrop KA, Phillips ES, Torrens C, Hanson MA, Jackson AA, Burdge GC: Feeding pregnant rats a protein-restricted diet persistently alters the methylation of specific cytosines in the hepatic PPAR alpha promoter of the offspring. British journal of nutrition. 2008; 100: 278–282. 30. Lillycrop KA, Slater-Jefferies JL, Hanson MA, Godfrey KM, Jackson AA, Burdge GC: Induction of altered epigenetic regulation of the hepatic glucocorticoid receptor in the offspring of rats fed a protein-restricted diet during pregnancy suggests that reduced DNA methyltransferase-1 expression is involved in impaired DNA methylation and changes in histone modifications. British journal of nutrition. 2007; 97: 1064–1073. 31. Pogribny IP, Starlard-Davenport A, Tryndyak VP, Han T, Ross SA, Rusyn I et al.: Difference in expression of hepatic microRNAs miR-29c, miR-34a, miR-155, and miR-200b is associated with strain-specific susceptibility to dietary nonalcoholic steatohepatitis in mice. Laboratory investigation. 2010; 90: 1437–1446. 32. Tryndyak VP, Ross SA, Beland FA, Pogribny IP: Down-regulation of the microRNAs miR-34a, miR-127, and miR-200b in rat liver during hepatocarcinogenesis induced by a methyl-deficient diet. Molecular carcinogenesis. 2009; 48: 479–487. 33. Marsit CJ, Eddy K, Kelsey KT: MicroRNA responses to cellular stress. Cancer research. 2006; 66: 10843–10848. 34. Cirera S, Birck M, Busk PK, Fredholm M: Expression profiles of miRNA-122 and its target CAT1 in minipigs (Sus scrofa) fed a high-cholesterol diet. Comparative medicine. 2010; 60: 136–141. 35. Salerno E, Scaglione BJ, Coffman FD, Brown BD, Baccarini A, Fernandes H et al.: Correcting miR-15a/16 genetic defect in New Zealand Black mouse model of CLL enhances drug sensitivity. Molecular cancer therapeutics. 2009; 8: 2684–2692. 36. Sun M, Estrov Z, Ji Y, Coombes KR, Harris DH, Kurzrock R: Curcumin (diferuloylmethane) alters the expression profiles of microRNAs in human pancreatic cancer cells. Molecular cancer therapeutics. 2008; 7: 464– 473. 37. Weiss FU, Marques IJ, Woltering JM, Vlecken DH, Aghdassi A, Partecke LI et al.: Retinoic acid receptor antagonists inhibit miR-10a expression and block metastatic behavior of pancreatic cancer. Gastroenterology. 2009; 137: 2136–2145.e1-7. 38. Martos SN, Tang WY, Wang Z: Elusive inheritance: Transgenerational effects and epigenetic inheritance in human environmental disease. Progress in biophysics and molecular biology. 2015; 118: 44–54. 39. Delage B, Dashwood RH: Dietary manipulation of histone structure and function. Annual review of nutrition. 2008; 28: 347–366.
NUTRIGENOMIK
3|2016 SZE 13