



Transkript
SERIE STEINZEITDIÄT
Essen wie in der Steinzeit – Darwin als ultimativer Ernährungsratgeber!?
Teil 4: Fragen der Evolutionsgenetik und präventivmedizinisches Potenzial der Paläodiät
ALEXANDER STRÖHLE* UND ANDREAS HAHN*
Alexander Ströhle
Im ersten Teil dieser Beitragsserie wurden die theoretischen Grundsäulen des Paläokonzepts dargestellt und die zugrunde liegende Anpassungsthese beleuchtet (1). Wie eine Ernährung nach dem «Paläoprinzip» beschaffen ist, wurde dann in den beiden letzten Beitragsteilen erläutert (2–3). In diesem abschliessenden Beitrag sollen die evolutionsgenetischen Annahmen des Paläokonzepts untersucht und eine ernährungsphysiologische und präventivmedizinische Bewertung vorgenommen werden.
Die These von der genetischen Konstanz auf dem Prüfstand
Zentral für das Paläokon-
Andreas Hahn
zept ist die Annahme, der Mensch habe im Verlauf
der letzten 10 000 Jahre
keine Möglichkeit gehabt, sich genetisch
an die «neuen» Lebens- und Ernährungs-
verhältnisse anzupassen. Denn, so das Ar-
gument, Evolution im Allgemeinen und
komplexe Anpassungsprozesse im Be-
sonderen nehmen sehr lange Zeiträume
in Anspruch (siehe Teil 1 dieser Beitrags-
serie; [1]). Was auf den ersten Blick plausi-
bel anmutet, hält einer Überprüfung bei
näherer Betrachtung aus mehreren Grün-
den nicht stand (5):
1. Genotypische versus phänotypische
Evolution. Ganz offensichtlich basiert das
Paläokonzept auf einer genzentrierten
Sicht von Evolution. Danach beruht evo-
lutionärer Wandel auf der Trias Mutation,
genetische Rekombination und Gen-
selektion. Dieser mikroreduktionistische
Ansatz wurde wiederholt kritisiert.
*Abteilung Ernährungsphysiologie und Humanernährung, Institut für Lebensmittelwissenschaft und Humanernährung, Leibniz Universität Hannover
Unbestritten ist, dass die genannten Mechanismen notwendige Elemente im Evolutionsgeschehen sind. Fraglich ist allerdings, ob sie hinreichend sind (6, 7). So fragte zum Beispiel bereits 1994 der an der Universität in Wien tätige Evolutionsbiologe Gerd B. Müller (8): «Ist die morpho-logische Evolution tatsächlich nur eine direkte Konsequenz der genetischen?
kann es zu starken Veränderungen kommen, ohne dass die Morphologie des Organismus davon nennenswert betroffen ist (Literatur bei [8, 9]). Entsprechend scheint auch die Geschwindigkeit der genetischen Veränderung (insbesondere der proteinkodierenden DNA-Sequenzen) im Verlauf der Evolution weitgehend konstant geblieben zu sein – unabhängig
«Human evolution didn’t stop when anatomically mo-
dern humans appeared, or when they expanded out of
»Africa. It never stopped – and why should it? (4)
Treibt (…) die kontinuierliche Modifikation des Genoms die morphologische Evolution in kleinen, graduellen Schritten voran und besteht der einzige richtungsgebende Faktor in der Selektion besser angepasster Phänotypen?» Heute ist klar, dass phänotypische Veränderungen nicht simplizistisch auf genotypische zu reduzieren sind. So belegen experimentelle Studien, dass die genetische und morphologische Evolution mitunter nur schwach korrelieren. Geringe Veränderungen auf Genebene können deutliche phänotypische Modifikationen bewirken und umgekehrt: Genotypisch
vom Tempo der morphologischen Evolution (10). Interessant sind in diesem Zusammenhang die Ergebnisse einer im Jahr 2007 publizierten Forschungsarbeit. Durch die Sequenzierung des Genoms der Spitzmausbeutelratte Monodelphis domestica wurde es möglich zu belegen, in welchem Ausmass es im Verlauf der Säugerevolution zu genetischen Veränderungen gekommen ist. Dabei zeigte sich: Obwohl die Aufspaltung der beiden Entwicklungslinien Metatheria (Beuteltiere) und Eutheria (Plazentatiere) etwa 180 Millionen Jahre zurückliegt, sind nur etwa 1 Prozent der proteinkodierenden
1 2/15
SERIE STEINZEITDIÄT
Genabschnitte spezifisch für höhere Säuger, das heisst, sie finden keine Entsprechung im SpitzmausbeutelrattenGenom (11). Auch ein Vergleich der orthologen kodierenden DNA-Sequenzen von Mensch und Schimpanse zeigt, dass die beiden Spezies zu 99 Prozent übereinstimmen, während bei 80 Prozent der Proteine Unterschiede bestehen (12) – von anatomischen und physiologischen Merkmalen ganz zu schweigen. Geringe genetische Unterschiede können also mit deutlichen phänotypischen – und damit auch potenziell funktionalen – Varianten korrespondieren, sodass die GenotypPhänotyp-Relation weit komplexer ist (13), als es von Anhängern der PaläoTheorie gemeinhin angenommen wird. 2. Die Geschwindigkeit evolutiven Wandels. Die Ansicht, die vergangenen 10 000 Jahre hätten für nennenswerte genetische Anpassungen zeitlich nicht ausgereicht, ignoriert jüngere Befunde der Evolutionsgenetik. So belegen etwa Feldstudien und experimentelle Untersuchungen, dass Artbildung (Speziation) und Populationsevolution mitunter sehr rasch erfolgen können (14). Ausgehend von 63 Studien wurde geschätzt, dass 25 Generationen ausreichen, damit sich ein Merkmal in einer Population via Selektion fixieren und ausbreiten kann (15). Auch eine bereits 1997 publizierte, am Süsswasserfisch Poecilia reticulata durchgeführte Untersuchung zeigte eindrucksvoll, wie rasch evolutionärer Wandel erfolgen kann. Gerade einmal 18 Generationen, so das Ergebnis der Untersuchung, sind notwendig, um spezifische
Lebenszyklusmerkmale auf Populationsebene abzuändern (16). Damit hätte seit Beginn der neolithischen Revolution ausreichend Zeit für relevante Anpassungsprozesse zur Verfügung gestanden (17, 18). Tatsächlich zeigen neueste Forschungsbefunde, dass sich das menschliche Genom gerade in der jüngeren Menschheitsgeschichte erheblich verändert hat ([19–28]; Übersicht bei [4, 29]). Dieser Umstand kann nicht verwundern, haben sich doch die Umweltbedingungen gerade im Verlauf des Holozäns in einem erheblichen Umfang verändert. Ob es sich um die Einführung des Ackerbaus, die Milchviehhaltung oder die höhere Bevölkerungsdichte handelt – von allen Faktoren dürfte in den letzten 10 000 Jahren ein erheblicher Selektionsdruck ausgegangen sein (29). In Summa lässt sich mit Blick auf die These von der genetischen Konstanz festhalten: Nur weil der Mensch 99 Prozent seiner Entwicklungsgeschichte als Jäger und Sammler und lediglich 1 Prozent unter neolithischen Bedingungen verbracht hat, bedeutet dies nicht, dass die letzten 6000 bis 10 000 Jahre evolutionsbiologisch irrelevant waren. Die Paläotheorie unterliegt hier offensichtlich einer Missinterpretation, die von dem US-Evolutionsbiologen David S. Wilson als «1-Prozent-Fehlschluss» bezeichnet wurde (18). Zudem stellt sich nicht die Frage, ob etwaige genetische Veränderungen in ihrem Ausmass gering ausgefallen sind oder nicht. Entscheidend ist lediglich, ob sie (ernährungs)physiologisch als relevant anzusehen sind (30).
Tabelle 1: Allgemeine nährstoffbezogene Charakterisierung der Jäger- und Sammler-Kost (31)
Nahrungsparameter Kohlenhydratanteile Fettanteil Proteinanteil Glykämische Last Gehalt an gesättigten Fettsäuren Gehalt an einfach ungesättigten Fettsäuren Gehalt an mehrfach ungesättigten Fettsäuren Gehalt an Omega-3-Fettsäuren Omega-3/Omega-6-Quotient Ballaststoffgehalt Natrium-Kalium-Verhältnis Säurelast
(Semi)quantitative Daten Niedrig bis moderat (20–40 Energie%) Niedrig bis moderat (28–47 Energie%) Hoch (19–35 Energie%) Niedrig Niedrig (7,5 Energie%) Hoch Moderat Hoch Hoch (2:1) Hoch (30 g/Tag) Niedrig (1:5) Basenüberschuss
Die Paläodiät im ernährungs-
physiologischen Test
Eine ernährungsphysiologische Bewertung ist dadurch erschwert, dass es sich bei der Steinzeit-Diät keinesfalls um eine homogene Ernährungsweise handelt. Vielmehr existieren verschiedene Ausprägungen, wobei die Empfehlungen zur Lebensmittelauswahl mitunter deutlich differieren. Dennoch lassen sich für die Paläodiät (ein mittleres Verhältnis vom Tier stammender und pflanzlicher Nahrungsmittel von etwa 1:1 vorausgesetzt) eine Reihe von allgemeinen Charakteristika identifizieren. Dazu zählen (31): • Makronährstoffverhältnis, worunter
die Energieprozentrelationen der Gesamtenergieaufnahme zu verstehen sind, die auf Kohlenhydrate, Fette und Proteine entfallen. • Glykämische Last (GL), das heisst die summierten und gemittelten Produkte aus glykämischem Index (GI) und dem Kohlenhydratgehalt (pro 100 g) aller verzehrten Lebensmittel. • Ballaststoffgehalt, also die absolute Masse an Ballaststoffen. • Fettsäurenzusammensetzung im Hinblick auf die absoluten und relativen Gehalte an gesättigten, einfach und mehrfach ungesättigten Fettsäuren sowie an Omega-3-, Omega-6- und Transfettsäuren. • Mikronährstoffdichte, das heisst der Quotient aus der Mineralstoff- beziehungsweise Vitaminmenge und dem Energiegehalt; meist angegeben in µg oder mg pro 1000 kcal oder 4,2 MJ. • Natrium-Kalium-Verhältnis • Säurelast, das heisst die von der Nahrung ausgehende Nettobelastung des Organismus mit fixen Säuren, ausgewiesen in Milliäquivalenten (mEq). Wie aus Tabelle 1 hervorgeht, weist die Paläodiät einen – im Vergleich mit den von nationalen Fachgesellschaften ausgesprochenen Empfehlungen – hohen Gehalt an Ballaststoffen, Vitaminen und Mineralstoffen auf. Bei Letzteren übersteigt der Kaliumgehalt den des Natriums um ein Vielfaches, sodass sich ein niedriger Natrium-Kalium-Quotient ergibt. Bei den Makronährstoffen imponiert der hohe Proteinanteil zu Lasten des Kohlen-
2/15
2
SERIE STEINZEITDIÄT
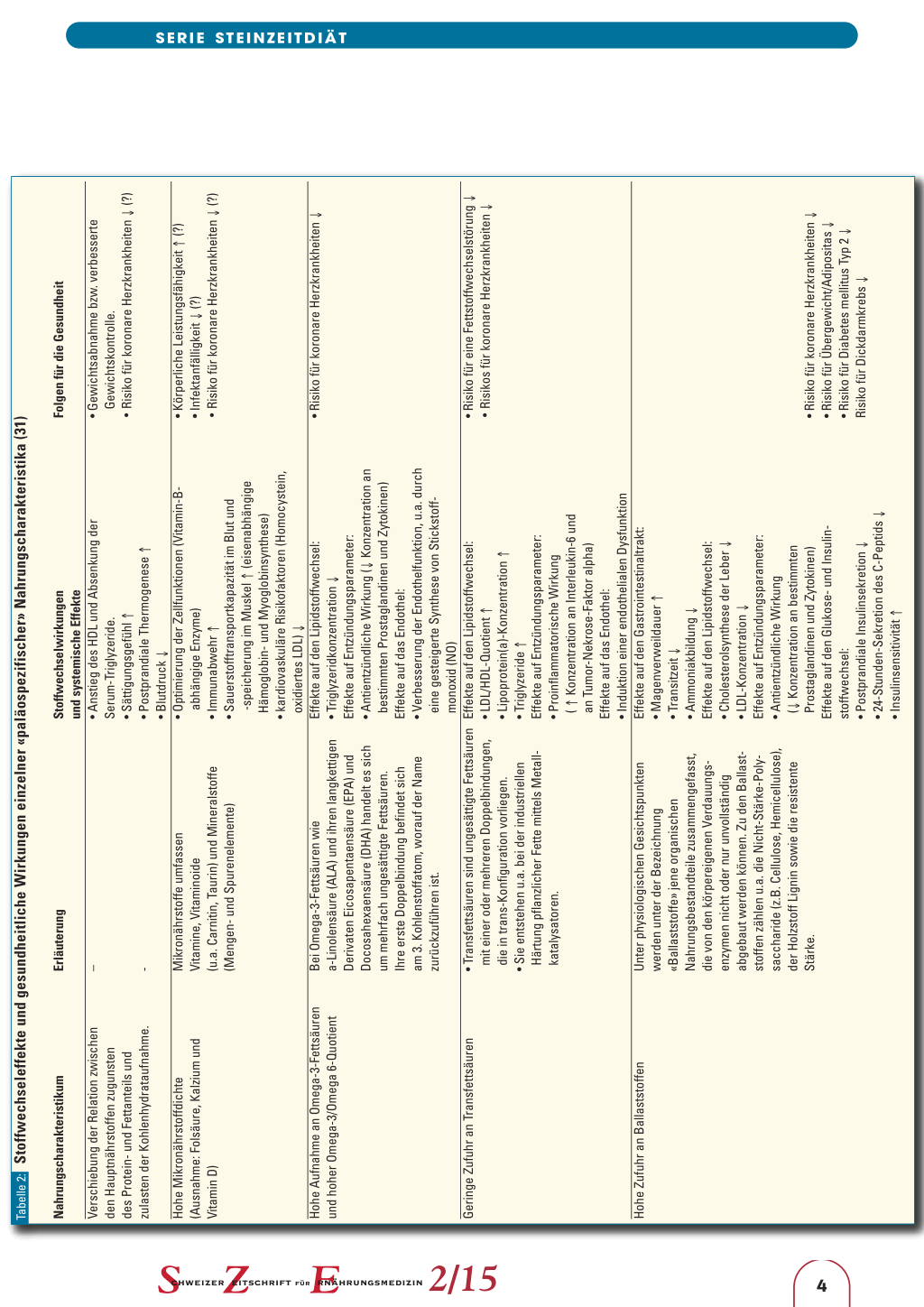
hydratanteils. Bedingt durch die hohe Aufnahme an Gemüsen, Wurzelknollen, Pilzen und Obst liefert die Kost einen Überschuss an Basenäquivalenten; auch die Zufuhr an sekundären Pflanzenstoffen dürfte hoch sein. Auf der anderen Seite führt der Verzicht auf Fertigprodukte, Erzeugnisse aus Auszugsmehlen und isolierten Zuckern zu einer niedrigen glykämischen Last; auch die Zufuhr an TransFettsäuren ist gering. Wie Tabelle 2 zeigt, sind die genannten Charakteristika der Paläo-Diät aus ernährungsphysiologischer Sicht insgesamt positiv zu beurteilen. Lediglich die hohe Fleischzufuhr und eine damit verbundene hohe Aufnahme an Hämeisen, heterozyklischen Aminen und polyzyklischen
Info 1: Stoffwechselstudien zur Paläodiät
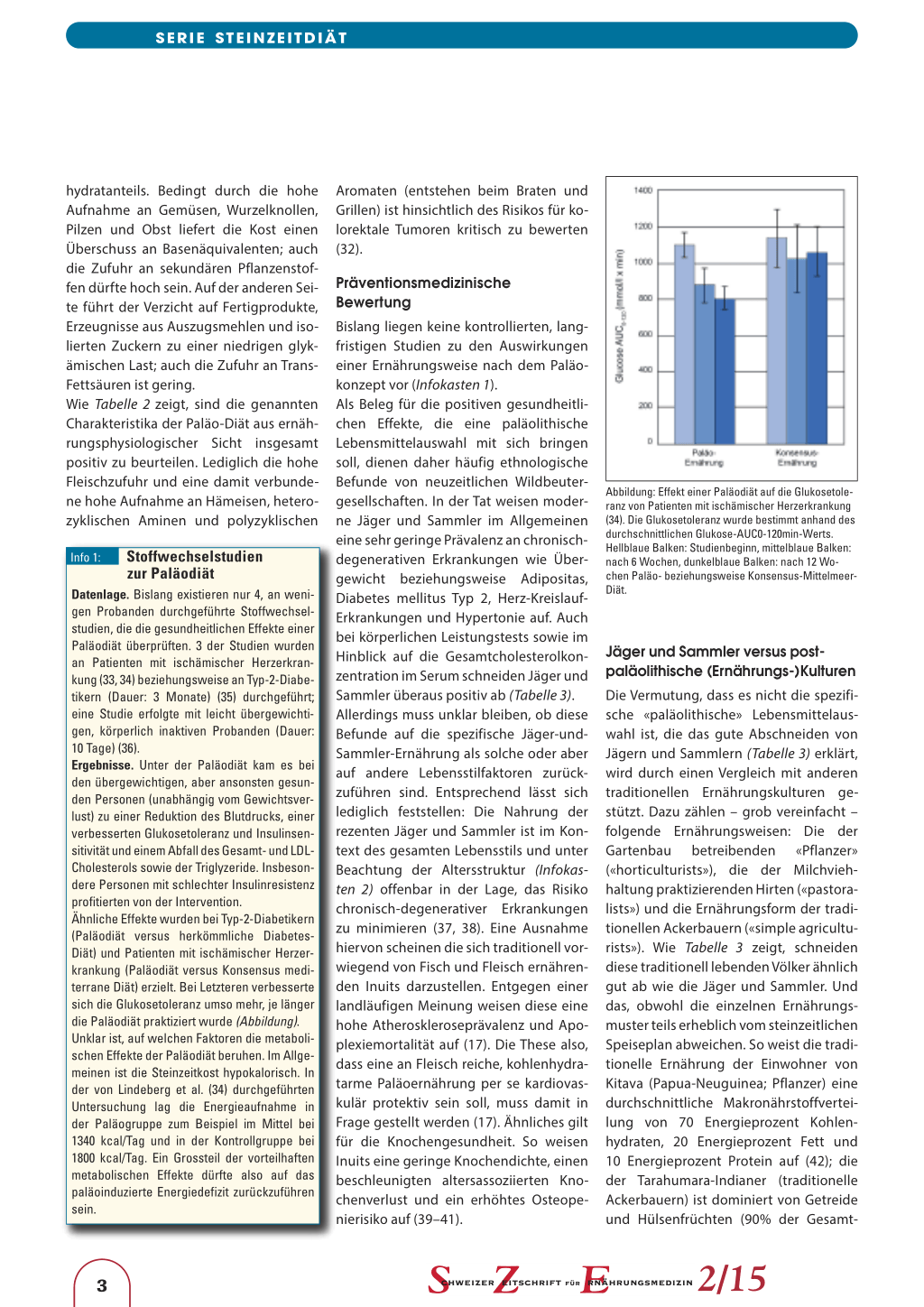
Datenlage. Bislang existieren nur 4, an wenigen Probanden durchgeführte Stoffwechselstudien, die die gesundheitlichen Effekte einer Paläodiät überprüften. 3 der Studien wurden an Patienten mit ischämischer Herzerkrankung (33, 34) beziehungsweise an Typ-2-Diabetikern (Dauer: 3 Monate) (35) durchgeführt; eine Studie erfolgte mit leicht übergewichtigen, körperlich inaktiven Probanden (Dauer: 10 Tage) (36). Ergebnisse. Unter der Paläodiät kam es bei den übergewichtigen, aber ansonsten gesunden Personen (unabhängig vom Gewichtsverlust) zu einer Reduktion des Blutdrucks, einer verbesserten Glukosetoleranz und Insulinsensitivität und einem Abfall des Gesamt- und LDLCholesterols sowie der Triglyzeride. Insbesondere Personen mit schlechter Insulinresistenz profitierten von der Intervention. Ähnliche Effekte wurden bei Typ-2-Diabetikern (Paläodiät versus herkömmliche DiabetesDiät) und Patienten mit ischämischer Herzerkrankung (Paläodiät versus Konsensus mediterrane Diät) erzielt. Bei Letzteren verbesserte sich die Glukosetoleranz umso mehr, je länger die Paläodiät praktiziert wurde (Abbildung). Unklar ist, auf welchen Faktoren die metabolischen Effekte der Paläodiät beruhen. Im Allgemeinen ist die Steinzeitkost hypokalorisch. In der von Lindeberg et al. (34) durchgeführten Untersuchung lag die Energieaufnahme in der Paläogruppe zum Beispiel im Mittel bei 1340 kcal/Tag und in der Kontrollgruppe bei 1800 kcal/Tag. Ein Grossteil der vorteilhaften metabolischen Effekte dürfte also auf das paläoinduzierte Energiedefizit zurückzuführen sein.
Aromaten (entstehen beim Braten und Grillen) ist hinsichtlich des Risikos für kolorektale Tumoren kritisch zu bewerten (32).
Präventionsmedizinische
Bewertung
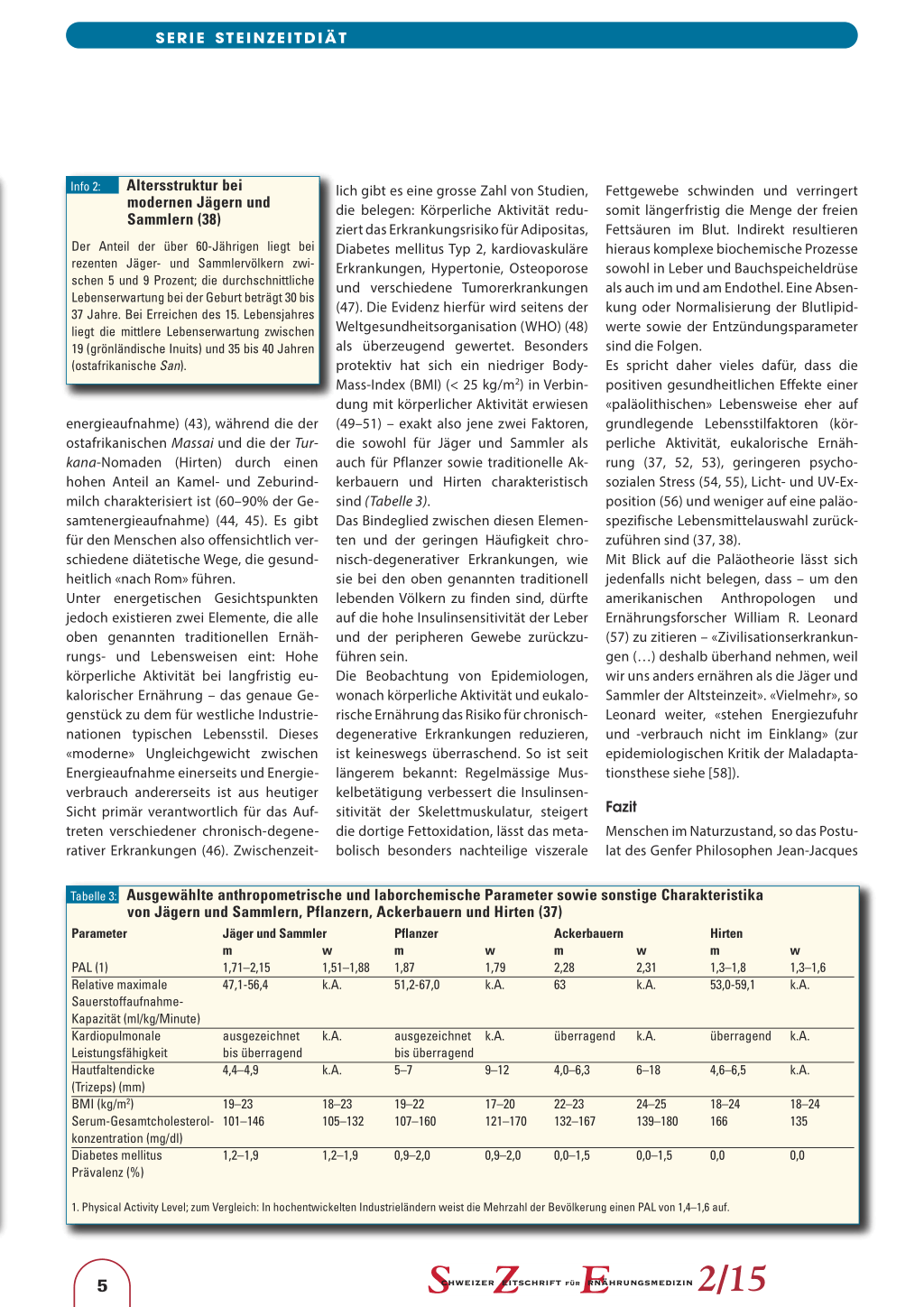
Bislang liegen keine kontrollierten, langfristigen Studien zu den Auswirkungen einer Ernährungsweise nach dem Paläokonzept vor (Infokasten 1). Als Beleg für die positiven gesundheitlichen Effekte, die eine paläolithische Lebensmittelauswahl mit sich bringen soll, dienen daher häufig ethnologische Befunde von neuzeitlichen Wildbeutergesellschaften. In der Tat weisen moderne Jäger und Sammler im Allgemeinen eine sehr geringe Prävalenz an chronischdegenerativen Erkrankungen wie Übergewicht beziehungsweise Adipositas, Diabetes mellitus Typ 2, Herz-KreislaufErkrankungen und Hypertonie auf. Auch bei körperlichen Leistungstests sowie im Hinblick auf die Gesamtcholesterolkonzentration im Serum schneiden Jäger und Sammler überaus positiv ab (Tabelle 3). Allerdings muss unklar bleiben, ob diese Befunde auf die spezifische Jäger-undSammler-Ernährung als solche oder aber auf andere Lebensstilfaktoren zurückzuführen sind. Entsprechend lässt sich lediglich feststellen: Die Nahrung der rezenten Jäger und Sammler ist im Kontext des gesamten Lebensstils und unter Beachtung der Altersstruktur (Infokasten 2) offenbar in der Lage, das Risiko chronisch-degenerativer Erkrankungen zu minimieren (37, 38). Eine Ausnahme hiervon scheinen die sich traditionell vorwiegend von Fisch und Fleisch ernährenden Inuits darzustellen. Entgegen einer landläufigen Meinung weisen diese eine hohe Atheroskleroseprävalenz und Apoplexiemortalität auf (17). Die These also, dass eine an Fleisch reiche, kohlenhydratarme Paläoernährung per se kardiovaskulär protektiv sein soll, muss damit in Frage gestellt werden (17). Ähnliches gilt für die Knochengesundheit. So weisen Inuits eine geringe Knochendichte, einen beschleunigten altersassoziierten Knochenverlust und ein erhöhtes Osteopenierisiko auf (39–41).
Abbildung: Effekt einer Paläodiät auf die Glukosetoleranz von Patienten mit ischämischer Herzerkrankung (34). Die Glukosetoleranz wurde bestimmt anhand des durchschnittlichen Glukose-AUC0-120min-Werts. Hellblaue Balken: Studienbeginn, mittelblaue Balken: nach 6 Wochen, dunkelblaue Balken: nach 12 Wochen Paläo- beziehungsweise Konsensus-MittelmeerDiät.
Jäger und Sammler versus postpaläolithische (Ernährungs-)Kulturen
Die Vermutung, dass es nicht die spezifische «paläolithische» Lebensmittelauswahl ist, die das gute Abschneiden von Jägern und Sammlern (Tabelle 3) erklärt, wird durch einen Vergleich mit anderen traditionellen Ernährungskulturen gestützt. Dazu zählen – grob vereinfacht – folgende Ernährungsweisen: Die der Gartenbau betreibenden «Pflanzer» («horticulturists»), die der Milchviehhaltung praktizierenden Hirten («pastoralists») und die Ernährungsform der traditionellen Ackerbauern («simple agriculturists»). Wie Tabelle 3 zeigt, schneiden diese traditionell lebenden Völker ähnlich gut ab wie die Jäger und Sammler. Und das, obwohl die einzelnen Ernährungsmuster teils erheblich vom steinzeitlichen Speiseplan abweichen. So weist die traditionelle Ernährung der Einwohner von Kitava (Papua-Neuguinea; Pflanzer) eine durchschnittliche Makronährstoffverteilung von 70 Energieprozent Kohlenhydraten, 20 Energieprozent Fett und 10 Energieprozent Protein auf (42); die der Tarahumara-Indianer (traditionelle Ackerbauern) ist dominiert von Getreide und Hülsenfrüchten (90% der Gesamt-
3 2/15
Tabelle 2: Stoffwechseleffekte und gesundheitliche Wirkungen einzelner «paläospezifischer» Nahrungscharakteristika (31)
SERIE STEINZEITDIÄT
2/15
Nahrungscharakteristikum Verschiebung der Relation zwischen den Hauptnährstoffen zugunsten des Protein- und Fettanteils und zulasten der Kohlenhydrataufnahme. Hohe Mikronährstoffdichte (Ausnahme: Folsäure, Kalzium und Vitamin D)
Hohe Aufnahme an Omega-3-Fettsäuren und hoher Omega-3/Omega 6-Quotient
Geringe Zufuhr an Transfettsäuren
Hohe Zufuhr an Ballaststoffen
Erläuterung
Stoffwechselwirkungen
und systemische Effekte
– • Anstieg des HDL und Absenkung der
Serum-Triglyzeride.
• Sättigungsgefühl ↑
- • Postprandiale Thermogenese ↑
• Blutdruck ↓
Mikronährstoffe umfassen
• Optimierung der Zellfunktionen (Vitamin-B-
Vitamine, Vitaminoide
abhängige Enzyme)
(u.a. Carnitin, Taurin) und Mineralstoffe
• Immunabwehr ↑
(Mengen- und Spurenelemente)
• Sauerstofftransportkapazität im Blut und
-speicherung im Muskel ↑ (eisenabhängige
Hämoglobin- und Myoglobinsynthese)
• kardiovaskuläre Risikofaktoren (Homocystein,
oxidiertes LDL) ↓
Bei Omega-3-Fettsäuren wie
Effekte auf den Lipidstoffwechsel:
a-Linolensäure (ALA) und ihren langkettigen • Triglyzeridkonzentration ↓
Derivaten Eicosapentaensäure (EPA) und
Effekte auf Entzündungsparameter:
Docosahexaensäure (DHA) handelt es sich • Antientzündliche Wirkung (↓ Konzentration an
um mehrfach ungesättigte Fettsäuren.
bestimmten Prostaglandinen und Zytokinen)
Ihre erste Doppelbindung befindet sich
Effekte auf das Endothel:
am 3. Kohlenstoffatom, worauf der Name
• Verbesserung der Endothelfunktion, u.a. durch
zurückzuführen ist.
eine gesteigerte Synthese von Stickstoff-
monoxid (NO)
• Transfettsäuren sind ungesättigte Fettsäuren Effekte auf den Lipidstoffwechsel:
mit einer oder mehreren Doppelbindungen, • LDL/HDL-Quotient ↑
die in trans-Konfiguration vorliegen.
• Lipoprotein(a)-Konzentration ↑
• Sie entstehen u.a. bei der industriellen
• Triglyzeride ↑
Härtung pflanzlicher Fette mittels Metall- Effekte auf Entzündungsparameter:
katalysatoren.
• Proinflammatorische Wirkung
( ↑ Konzentration an Interleukin-6 und
an Tumor-Nekrose-Faktor alpha)
Effekte auf das Endothel:
• Induktion einer endothelialen Dysfunktion
Unter physiologischen Gesichtspunkten
Effekte auf den Gastrointestinaltrakt:
werden unter der Bezeichnung
• Magenverweildauer ↑
«Ballaststoffe» jene organischen
• Transitzeit ↓
Nahrungsbestandteile zusammengefasst,
• Ammoniakbildung ↓
die von den körpereigenen Verdauungs-
Effekte auf den Lipidstoffwechsel:
enzymen nicht oder nur unvollständig
• Cholesterolsynthese der Leber ↓
abgebaut werden können. Zu den Ballast- • LDL-Konzentration ↓
stoffen zählen u.a. die Nicht-Stärke-Poly-
Effekte auf Entzündungsparameter:
saccharide (z.B. Cellulose, Hemicellulose), • Antientzündliche Wirkung
der Holzstoff Lignin sowie die resistente
(↓ Konzentration an bestimmten
Stärke.
Prostaglandinen und Zytokinen)
Effekte auf den Glukose- und Insulin-
stoffwechsel:
• Postprandiale Insulinsekretion ↓
• 24-Stunden-Sekretion des C-Peptids ↓
• Insulinsensitivität ↑
Folgen für die Gesundheit • Gewichtsabnahme bzw. verbesserte
Gewichtskontrolle. • Risiko für koronare Herzkrankheiten ↓ (?) • Körperliche Leistungsfähigkeit ↑ (?) • Infektanfälligkeit ↓ (?) • Risiko für koronare Herzkrankheiten ↓ (?)
• Risiko für koronare Herzkrankheiten ↓
• Risiko für eine Fettstoffwechselstörung ↓ • Risikos für koronare Herzkrankheiten ↓
• Risiko für koronare Herzkrankheiten ↓ • Risiko für Übergewicht/Adipositas ↓ • Risiko für Diabetes mellitus Typ 2 ↓ Risiko für Dickdarmkrebs ↓
4
SERIE STEINZEITDIÄT
Info 2:
Altersstruktur bei modernen Jägern und Sammlern (38)
Der Anteil der über 60-Jährigen liegt bei rezenten Jäger- und Sammlervölkern zwischen 5 und 9 Prozent; die durchschnittliche Lebenserwartung bei der Geburt beträgt 30 bis 37 Jahre. Bei Erreichen des 15. Lebensjahres liegt die mittlere Lebenserwartung zwischen 19 (grönländische Inuits) und 35 bis 40 Jahren (ostafrikanische San).
energieaufnahme) (43), während die der ostafrikanischen Massai und die der Turkana-Nomaden (Hirten) durch einen hohen Anteil an Kamel- und Zeburindmilch charakterisiert ist (60–90% der Gesamtenergieaufnahme) (44, 45). Es gibt für den Menschen also offensichtlich verschiedene diätetische Wege, die gesundheitlich «nach Rom» führen. Unter energetischen Gesichtspunkten jedoch existieren zwei Elemente, die alle oben genannten traditionellen Ernährungs- und Lebensweisen eint: Hohe körperliche Aktivität bei langfristig eukalorischer Ernährung – das genaue Gegenstück zu dem für westliche Industrienationen typischen Lebensstil. Dieses «moderne» Ungleichgewicht zwischen Energieaufnahme einerseits und Energieverbrauch andererseits ist aus heutiger Sicht primär verantwortlich für das Auftreten verschiedener chronisch-degenerativer Erkrankungen (46). Zwischenzeit-
lich gibt es eine grosse Zahl von Studien, die belegen: Körperliche Aktivität reduziert das Erkrankungsrisiko für Adipositas, Diabetes mellitus Typ 2, kardiovaskuläre Erkrankungen, Hypertonie, Osteoporose und verschiedene Tumorerkrankungen (47). Die Evidenz hierfür wird seitens der Weltgesundheitsorganisation (WHO) (48) als überzeugend gewertet. Besonders protektiv hat sich ein niedriger BodyMass-Index (BMI) (< 25 kg/m2) in Verbindung mit körperlicher Aktivität erwiesen (49–51) – exakt also jene zwei Faktoren, die sowohl für Jäger und Sammler als auch für Pflanzer sowie traditionelle Akkerbauern und Hirten charakteristisch sind (Tabelle 3). Das Bindeglied zwischen diesen Elementen und der geringen Häufigkeit chronisch-degenerativer Erkrankungen, wie sie bei den oben genannten traditionell lebenden Völkern zu finden sind, dürfte auf die hohe Insulinsensitivität der Leber und der peripheren Gewebe zurückzuführen sein. Die Beobachtung von Epidemiologen, wonach körperliche Aktivität und eukalorische Ernährung das Risiko für chronischdegenerative Erkrankungen reduzieren, ist keineswegs überraschend. So ist seit längerem bekannt: Regelmässige Muskelbetätigung verbessert die Insulinsensitivität der Skelettmuskulatur, steigert die dortige Fettoxidation, lässt das metabolisch besonders nachteilige viszerale Fettgewebe schwinden und verringert somit längerfristig die Menge der freien Fettsäuren im Blut. Indirekt resultieren hieraus komplexe biochemische Prozesse sowohl in Leber und Bauchspeicheldrüse als auch im und am Endothel. Eine Absenkung oder Normalisierung der Blutlipidwerte sowie der Entzündungsparameter sind die Folgen. Es spricht daher vieles dafür, dass die positiven gesundheitlichen Effekte einer «paläolithischen» Lebensweise eher auf grundlegende Lebensstilfaktoren (körperliche Aktivität, eukalorische Ernährung (37, 52, 53), geringeren psychosozialen Stress (54, 55), Licht- und UV-Exposition (56) und weniger auf eine paläospezifische Lebensmittelauswahl zurückzuführen sind (37, 38). Mit Blick auf die Paläotheorie lässt sich jedenfalls nicht belegen, dass – um den amerikanischen Anthropologen und Ernährungsforscher William R. Leonard (57) zu zitieren – «Zivilisationserkrankungen (…) deshalb überhand nehmen, weil wir uns anders ernähren als die Jäger und Sammler der Altsteinzeit». «Vielmehr», so Leonard weiter, «stehen Energiezufuhr und -verbrauch nicht im Einklang» (zur epidemiologischen Kritik der Maladaptationsthese siehe [58]). Fazit Menschen im Naturzustand, so das Postulat des Genfer Philosophen Jean-Jacques Tabelle 3: Ausgewählte anthropometrische und laborchemische Parameter sowie sonstige Charakteristika von Jägern und Sammlern, Pflanzern, Ackerbauern und Hirten (37) Parameter Jäger und Sammler mw PAL (1) 1,71–2,15 1,51–1,88 Relative maximale 47,1-56,4 k.A. Sauerstoffaufnahme- Kapazität (ml/kg/Minute) Kardiopulmonale ausgezeichnet k.A. Leistungsfähigkeit bis überragend Hautfaltendicke 4,4–4,9 k.A. (Trizeps) (mm) BMI (kg/m2) 19–23 18–23 Serum-Gesamtcholesterol- 101–146 105–132 konzentration (mg/dl) Diabetes mellitus 1,2–1,9 1,2–1,9 Prävalenz (%) Pflanzer m 1,87 51,2-67,0 w 1,79 k.A. ausgezeichnet k.A. bis überragend 5–7 9–12 19–22 107–160 17–20 121–170 0,9–2,0 0,9–2,0 Ackerbauern m 2,28 63 w 2,31 k.A. überragend k.A. 4,0–6,3 6–18 22–23 132–167 24–25 139–180 0,0–1,5 0,0–1,5 Hirten m 1,3–1,8 53,0-59,1 überragend 4,6–6,5 18–24 166 0,0 w 1,3–1,6 k.A. k.A. k.A. 18–24 135 0,0 1. Physical Activity Level; zum Vergleich: In hochentwickelten Industrieländern weist die Mehrzahl der Bevölkerung einen PAL von 1,4–1,6 auf. 5 2/15 SERIE STEINZEITDIÄT Rousseau, seien «kräftig, leichtfüssig und klaren Auges». Wenngleich Anthropologen sich mitunter skeptisch zu den Lebensbedingungen und zum Gesundheitszustand unserer altseinzeitlichen Vorfahren äussern (59–61), so ist offenkundig: «im Naturzustand» sind chronisch-degenerative Erkrankungen wie Adipositas, essenzielle Hypertonie, koronare Herzerkrankungen und Diabetes mellitus Typ 2 selten zu beobachten (47, 62). Die Ursachen hierfür sind vielschichtig. Neben körperlicher Aktivität, einer kalorisch knappen, aber mikronährstoff- sowie ballaststoffreichen Kost und einer damit verbundenen schlanken Körperstatur dürften auch psychosoziale (kleine Gruppen, Fehlen von chronischem Dystress) und chronobiologische Faktoren (ausreichend Schlaf ) von gesundheitlicher Relevanz sein. Zurück also in die Steinzeit? Vielleicht partiell. Aber sicher nicht in Gänze für das Gebiet der Ernährung. In der Tat gibt es keinen wissenschaftlichen Grund, weshalb man kulturhistorisch zu einer altsteinzeitlichen Lebensmittelauswahl zurückkehren und die Errungenschaften traditioneller Ernährungskulturen ernährungsphysiologisch diffamieren müsste. Sollte die Botschaft einer modernen, liberalisierten Paläodiät jedoch lauten, bei der Nahrungsauswahl Obst, Gemüse, Hülsenfrüchte, Nüsse, mageres Fleisch und Fisch zu präferieren, «Junk Food» möglichst zu meiden (63, 64) und Vollkorn- sowie Milchprodukte in Abhängigkeit von der individuellen Verträglichkeit zu konsumieren (65) – dann ist eine solche Empfehlung nur zu begrüssen. Korrespondenzadresse: Dr. Alexander Ströhle Leibniz Universität Hannover Institut für Lebensmittelwissenschaft und Humanernährung Abteilung Ernährungsphysiologie und Humanernährung Am Kleinen Felde 30 D-30167 Hannover E-Mail: alexander.stroehle@gmx.de Literatur: 1. Ströhle A, Hahn A. Essen wie in der Steinzeit – Darwin als ultimativer Ernährungsratgeber? Teil 1: Vom Paläokonzept zum Anpassungsparadigma. Schweiz Z Ernährungsmed. 2014; 2: 41–49. 2. Ströhle A, Hahn A. Essen wie in der Steinzeit – Darwin als ultimativer Ernährungsratgeber? Teil 2: Auf der Suche nach «der» paläolithischen Ernährung - paläoökologische Befunde. Schweiz Z Ernährungsmed 2014; 33–41. 3. Ströhle A, Hahn A. Essen wie in der Steinzeit – Darwin als ultimativer Ernährungsratgeber? Teil 3: Auf der Suche nach «der» paläolithischen Ernährung - Ethnologische Befunde. Schweiz Z Ernährungsmed 2014; 5: 34–38. 4. Cochran G, Harpending H: The 10,000 year explosion. How civilization accelerated human evolution. Basic Books, New York 2009. 5. Ströhle A. Der moderne Mensch, ein Gefangener des Paläolithikums? Eine kritische Analyse der evolutionsmedizinischen Mismatch-Theorie. Naturw Rdsch. 2012; 65: 1–13. 6. Wagner GP, Laubichler MD. Rupert Riedl and the resynthesis of evolutionary and developmental biology: body plans and evolvability. J Exp Zoolog B Mol Dev Evol 2004; 302: 92–102. 7. Wagner GP. What is the promise of developmental evolution? Part I: why is developmental biology necessary to explain evolutionary innovations? J Exp Zool 2000; 288: 95–98. 8. Müller GB. Evolutionäre Entwicklungsbiologie: Grundlage einer neuen Synthese. In: Wieser W (Hrsg.): Die Evolution der Evolutionstheorie. Von Darwin zur DNA. Wissenschaftliche Buchgesellschaft, Darmstadt 1994, S. 154–193. 9. Newman SA, Müller GB. Epigenetic mechanisms of character origination. J Exp Zool Mol Dev Evol 2000; 288: 304–317. 10. Maier UG. Die molekulare Dimension der Evolution. In: Wieser W (Hrsg.): Die Evolution der Evolutionstheorie. Von Darwin zur DNA. Wissenschaftliche Buchgesellschaft, Darmstadt 1994, 109–128. 11. Mikkelsen TS, Wakefield MJ, Aken B, Amemiya CT, Chang JL, Duke S, Garber M, Gentles AJ, Goodstadt L, Heger A, Jurka J, Kamal M, Mauceli E, Searle SM, Sharpe T, Baker ML, Batzer MA, Benos PV, Belov K, Clamp M, Cook A, Cuff J, Das R,et al. Genome of the marsupial Monodelphis domestica reveals innovation in non-coding sequences. Nature. 2007; 447 (7141): 167–177. 12. Glazko G, Veeramachaneni V, Nei M, Makalowski W. Eighty percent of proteins are different between humans and chimpanzees. Gene 2005; 346: 215–219. 13. Moss L. Redundancy, plasticity, and detachment: The implications of comparative genomics for evolutionary thinking. Philos Sci 2006; 73: 930–946. 14. Thompson JN. Rapid evolution as ecological process. Trends Ecol Evol 1998; 13: 329–332. 15. Kingsolver JG, Hoekstra HE, Hoekstra JM, Berrigan D, Vignieri SN, Hill CE, Hoang A, Gilbert P. The strength of phenotypic selection in natural populations. Am Nat 2001; 157: 245–261. 16. Reznick DN, Shaw FH, Rodd FH, Shaw RG. Evaluation of the Rate of Evolution in Natural Populations of Guppies (Poecilia reticulata). Science 1997; 275 (5308): 1934–1937. 17. Ströhle A, Wolters M, Hahn A. Carbohydrates and the diet-atherosclerosis connection – More between earth and heaven. Comment on the article «The atherogenic potential of dietary carbohydrate». Prev Med 2007; 44: 82–84. 18. Wilson DS. Adaptive Genetic variation and human evolutionary psychology. Ethol Sociobiol 1004; 15: 219–235. 19. Lappalainen T, Salmela E, Andersen PM, DahlmanWright K, Sistonen P, Savontaus ML, Schreiber S, Lahermo P, Kere J. Genomic landscape of positive natural selection in Northern European populations. Eur J Hum Genet. 2010;18 (4): 471–478. 20. Colonna V, Ayub Q, Chen Y, Pagani L, Luisi P, Pybus M, Garrison E, Xue Y, Tyler-Smith C; 1000 Genomes Project Consortium, Abecasis GR, Auton A, Brooks LD, DePristo MA, Durbin RM, Handsaker RE, Kang HM, Marth GT, McVean GA. Human genomic regions with exceptionally high levels of population differentiation identified from 911 whole-genome sequences. Genome Biol 2014; 15: R88. 21. Richerson PJ, Boyd R, Henrich J. Colloquium paper: gene-culture coevolution in the age of genomics. Proc Natl Acad Sci 2010; 107 Suppl 2: 8985–8992. 22. Hawks J, Wang ET, Cochran GM, Harpending HC, Moyzis RK. Recent acceleration of human adaptive evolution. Proc Natl Acad Sci 2007; 104: 20753–20758. 23. Gibbons A. Human Evolution. Tracing evolution’s recent fingerprints. Science 2010; 329 (5993): 740–742. 24. Kuan V, Martineau AR, Griffiths CJ, Hyppönen E, Walton R. DHCR7 mutations linked to higher vitamin D status allowed early human migration to northern latitudes. BMC Evol Biol 2013; 13: 144. 25. Ségurel L, Austerlitz F, Toupance B, Gautier M, Kelley JL, Pasquet P, Lonjou C, Georges M, Voisin S, Cruaud C, Couloux A, Hegay T, Aldashev A, Vitalis R, Heyer E. Positive selection of protective variants for type 2 diabetes from the Neolithic onward: a case study in Central Asia. Eur J Hum Genet 2013; 10: 1146–1151. 26. Schlebusch CM, Sjödin P, Skoglund P, Jakobsson M. Stronger signal of recent selection for lactase persistence in Maasai than in Europeans. Eur J Hum Genet 2013; 21: 550–553. 27. Prendergast JG, Semple CA. Widespread signatures of recent selection linked to nucleosome positioning in the human lineage. Genome Res 2011; 21: 1777–1787. 28. Klimentidis YC, Abrams M, Wang J, Fernandez JR, Allison DB. Natural selection at genomic regions associated with obesity and type-2 diabetes: East Asians and sub-Saharan Africans exhibit high levels of differentiation at type-2 diabetes regions. Hum Genet. 2011; 129: 407–418. 29. Laland KN, Odling-Smee J, Myles S. How culture shaped the human genome: bringing genetics and the human sciences together. Nat Rev Genet 2010; 11: 137–148. 30. Buller DJ. Evolutionary Psychology: A Critique. In: Sober E (ed.): Conceptual Issues in Evolutionary Biology. 3rd ed., MIT press, Cambridge/Massachusetts/ London 2006, pp. 197–214. 31. Ströhle A, Hahn A. Ernährung á la Altsteinzeit – Ultima Ratio der Prävention? Dtsch Apothek Z. 2011; 151: 54–62. 32. Corpet DE. Red meat and colon cancer: should we become vegetarians, or can we make meat safer? Meat Sci. 2011; 89: 310–316. 33. Jönsson T, Granfeldt Y, Erlanson-Albertsson C, Ahrén B, Lindeberg S. A paleolithic diet is more satiating per calorie than a mediterranean-like diet in individuals with ischemic heart disease. Nutr Metab (Lond) 2010; 7:85. 34. Lindeberg S, Jönsson T, Granfeldt Y, Borgstrand E, Soffman J, Sjöström K, Ahrén B. A Palaeolithic diet improves glucose tolerance more than a Mediterranean-like diet in individuals with ischaemic heart disease. Diabetologia 2007; 50: 1795–1807. 35. Jönsson T, Granfeldt Y, Ahrén B, Branell UC, Pålsson G, Hansson A, Söderström M, Lindeberg S. Beneficial effects of a Paleolithic diet on cardiovascular risk factors in type 2 diabetes: a randomized crossover pilot study. Cardiovasc Diabetol 2009; 8: 35. 36. Frassetto LA, Schloetter M, Mietus-Synder M, Morris RC Jr, Sebastian A. Metabolic and physiologic improvements from consuming a paleolithic, huntergatherer type diet. Eur J Clin Nutr 2009; 63: 947–955. 2/15 6 SERIE STEINZEITDIÄT 37. Ströhle A, Wolters M, Hahn A. Die Ernährung des Menschen im evolutionsmedizinischen Kontext, Wien Klin Wochenschr 2009; 121: 173–187. 38. Ströhle A. Sub specie evolutionis. Eine Studie zur Evolutionären Ernährungswissenschaft, Shaker, Aachen 2010. 39. Mazess RB, Mather WE. Bone mineral content in Canadian Eskimos. Hum Biol 1975; 47: 44–63. 40. Mazess RB, Mather W. Bone mineral content of North Alaskan Eskimos. Am J Clin Nutr 1974; 27: 916–925. 41. Mazess RB. Bone density in Sadlermiut Eskimo. Hum Biol 1966; 38: 42–49. 42. Lindeberg S, Apparent absence of cerebrocardiovascular disease in Melanesians. Risk factors and nutritional considerations – the Kitava Study. University of Lund, Lund (1994). 43. Cerqueira MT, Fry MM, Connor WE, The food and nutrient intakes of the Tarahumara Indians of Mexico. Am J Clin Nutr 1979; 32, 905–915. 44. Ho KJ, The Massai of east africa: Some Unique biological characteristics. Arch Path 1971; 91: 387–410. 45. Little MA, Human biology, health and ecology of nomadic Turkana pastoralis. In: Leonard WR, Crawford MH (eds.): Human Biology of Pastoral Populations. Cambridge University Press, Cambridge (2002) pp. 151–182. 46. Ströhle A, Hahn A. Was Evolution nicht lehrt. Ernähr-Umschau 2003; 50: 481–482. 47. Ströhle A. Rück- und Seitenblicke im Zeitalter der Ernährungsver(w)irrung – Eine Hommage an Werner Kollath., Reglin Verlag, Köln 2009. 48. WHO/FAO. Diet, Nutrition and the Prevention of Chronic Diseases. Report of a Joint WHO/FAO Expert Consultation. Technical Report Series No. 916, Geneva 2003. 49. Rana JS, Li TY, Manson JE, Hu FB. Adiposity compared with physical inactivity nd risk of type 2 diabetes in women. Diabetes Care. 2007; 30: 53–58. 50. Li TY, Rana JS, Manson JE, Willett WC, Stampfer MJ, Colditz GA, Rexrode KM, Hu FB. Obesity as compared with physical activity in predicting risk of coronary heart disease in women. Circulation. 2006; 113: 499–506. 51. Hu FB, Willett WC, Li T, Stampfer MJ, Colditz GA, Manson JE. Adiposity as compared with physical activity in predicting mortality among women. N Engl J Med. 2004; 351: 2694–2703. 52. Gutin B, Owens S. The influence of physical activity on cardiometabolic biomarkers in youths: a review. Pediatr Exerc Sci 2011; 23: 169–185. 53. Archer E, Blair SN. Physical activity and the prevention of cardiovascular disease: from evolution to epidemiology. Prog Cardiovasc Dis 2011; 53: 387–396. 54. Gundersen C, Mahatmya D, Garasky S, Lohman B. Linking psychosocial stressors and childhood obesity. Obes Rev 2011; 12: e54-63. 55. Low CA, Thurston RC, Matthews KA. Psychosocial factors in the development of heart disease in women: current research and future directions. Psychosom Med 2010; 72: 842–854. 56. Grant WB, Juzeniene A, Moan JE. Review Article: Health benefit of increased serum 25(OH)D levels from oral intake and ultraviolet-B irradiance in the Nordic countries. Scand J Public Health 2011; 39: 70. 57. Leonard WR. Menschwerdung durch Kraftnahrung. In: N.N. Die Evolution des Menschen II. Spektrum der Wissenschaft Dossier 1/2004. 58. Gage TB. Are modern environments really bad for us? Revisting the demographic and epidemiologic transitions. Yrbk Phys Anthropol 2005; 48: 96–117. 59. Bogin B. !Kung nutritional status and the original «affluent society» – a new analysis. Anthropol Anz 2011; 68: 349–366. 60. Clarke EM, Thompson RC, Allam AH, Wann LS, Lombardi GP, Sutherland ML, Sutherland JD, Cox SL, Soliman MA, Abd el-Maksoud G, Badr I, Miyamoto MI, Frohlich B, Nur el-din AH, Stewart AF, Narula J, Zink AR, Finch CE, Michalik DE, Thomas GS. Is atherosclerosis fundamental to human aging? Lessons from ancient mummies. J Cardiol 2014; 63: 329–334. 61. Thompson RC, Allam AH, Lombardi GP, Wann LS, Sutherland ML, Sutherland JD, Soliman MA, Frohlich B, Mininberg DT, Monge JM, Vallodolid CM, Cox SL, Abd el-Maksoud G, Badr I, Miyamoto MI, el-Halim Nur el-Din A, Narula J, Finch CE, Thomas GS. Atherosclerosis across 4000 years of human history: the Horus study of four ancient populations. Lancet. 2013; 381: 1211–1222. 62. Eaton SB, Konner M, Shostak M. Stone agers in the fast lane: chronic degenerative diseases in evolutionary perspective. Am J Med 1988; 84: 739–749. 63. O’Keefe JH Jr, Cordain L. Cardiovascular disease resulting from a diet and lifestyle at odds with our Paleolithic genome: how to become a 21st-century hunter-gatherer. Mayo Clin Proc 2004; 79: 101–108. 64. Lindeberg S, Cordain L, Eaton SB. Biological and clinical potential of a paleolithic diet. J Nutr Environ Med 2003; 13: 149–160. 65. Junker T, Paul S: Der Darwin Code. Die Evolution erklärt unser Leben. C.H. Beck, Beck’sche Reihe, München 2010. 7 2/15