


Transkript
Entzündung und Ernährung
Der Darm als Immunorgan
Die Bedeutung der Darmflora für die Gesundheit
Ein intaktes und reguliertes intestinales
Immunsystem ist entscheidend für
einen gesunden Darm. Das Immun-
system des Darms ist sehr komplex
aufgebaut und eng verbunden mit der
Darmflora und dem Epithel. Für den
Organismus ist es lebensnotwendig,
dass dieses System pathogene Keime
rechtzeitig erkennt und unschädlich
macht. Viele Darmerkrankungen las-
sen sich auf eine gestörte Darmflora
und eine defekte Darmbarriere zurück-
führen. Prä- und Probiotika sind in die-
sem Zusammenspiel sehr interessant,
da sie im Darm eine Reihe nützlicher
Wirkungen haben. Einige Keime wer-
den inzwischen bereits erfolgreich zur
Prävention und Therapie intestinaler
Erkrankungen eingesetzt.
Rémy Meier
Darmflora
Die Gesundheit des Menschen hängt eng mit einer ausgewogenen Darmflora, einer intakten Darmbarriere und einem regulierten Immunsystem zusammen. Im menschlichen Dickdarm befinden sich etwa 0,5 bis 1 kg Bakterien. Die Anzahl der Bakterien ist mit 1013 bis 1014 zehnmal grösser als die Gesamtheit der Körperzellen. Von den über 500 verschiedenen im Gastrointestinaltrakt lebenden Keimen sind die meisten anaerob, das heisst, sie benötigen keinen freien Sauerstoff
zum Leben. Zwischen Darmflora und Wirt besteht eine Lebensgemeinschaft mit gegenseitigem Nutzen.
Die Zusammensetzung der Darmflora ist individuell, unterscheidet sich also von Mensch zu Mensch. Nach dem dritten Lebensjahr bleibt sie relativ konstant und verändert sich erst wieder im Alter. Die Darmflora enthält ein Gemisch apathogener und pathogener Keime; solange diese beiden Populationen im Gleichgewicht sind, ist der Darm gesund (1, 2). Um dieses Gleichgewicht zu gewährleisten, kontrolliert sich die Darmflora teilweise selbst, indem verschiedene Keime sogenannte Bakteriozine (antibakterielle Proteine) sezernieren, die das Wachstum pathogener Keime hemmen können (3). Die Panethzellen des Darmepithels bilden zudem Defensine, die – ähnlich wie die Bakteriozine – das Überwuchern von pathogenen Keimen kontrollieren.
Die Darmbakterien sind äusserst aktiv und beeinflussen metabolische, trophische und protektive Funktionen. Darüber hinaus fermentieren sie unverdaute in den Dickdarm gelangte Kohlenhydrate. Daraus entstehen sehr aktive kurzkettige Fettsäuren (KKFS) wie Butyrat, Propionat und Acetat. Das Dickdarmepithel bezieht seine notwendige Energie zu 80 Prozent von dem im Darm gebildeten Butyrat. Die KKFS stimulieren das Epithelwachstum und seine Differenzierung und sind an der Natrium- und Wasserresorption beteiligt. Im Weiteren hat Butyrat antientzündliche Eigenschaften. Es hemmt nicht nur den intrazellulären Transkriptionsfaktor NF-κB, sondern zeigt darüber hinaus auch einen protektiven dualen Effekt: Es reduziert proinflammatorische Zytokine (Interleukin 1, Interleukin 6, TNF-α) und kann antiinflammatorische Zytokine wie Interleukin 10 stimulieren (4, 5).
Darmepithel
Das mit einer Mukusschicht überzogene Epithel hat im Darm eine wichtige Barrierefunktion. Es ist bekannt, dass apathogene Bakterien zusammen mit dem Mukus die Darmbarriere verstärken und so die Aufnahme pathogener Keime und Fremdpartikel verhin-
dern. Das Epithel besteht aus einer einlagigen Zellschicht. Spezielle Kittleisten (tight junctions) verbinden die Epithelzellen untereinander. Dies ist ein weiterer Schutz gegen das Eindringen von Mikroorganismen. Die Epithelzellen sind nicht uniform, einige davon haben sehr spezifische Aufgaben (6). Wichtig zu erwähnen sind die endokrinen Zellen, die sogenannten Panethzellen (Defensinproduktion) und die Becherzellen (Schleimproduktion). Normalerweise können keine Bakterien durch das intakte Darmepithel hindurch diffundieren. Intestinale Bakterien können aber mithilfe spezialisierter Zellen (M-Zellen, dendritische Zellen) mit dem intestinalen Immunsystem kommunizieren. Die MZellen (microfold cells) sind ins Epithel eingestreut und liegen vor allem über den peyerschen Platten. Mit diesen M-Zellen können pathogene Bakterien direkt aufgenommen und inaktiviert werden (7). Durch das Epithel ziehen auch die Ausläufer der dendritischen Zellen, die Antigene aus dem Darmlumen aufnehmen und so in den peyerschen Platten (durch die Antigenpräsentation) T- und B-Lymphozyten aktivieren können.
Im Epithel sind auch viele T-Lymphozyten angesiedelt. Diese sind wichtig für die erste Abwehr von Bakterien und Viren. Sie können sowohl zytotoxisch aktiv sein als auch als Helferzellen dienen. Die Darmflora, das Darmepithel und das intestinale Immunsystem sind eng miteinander verknüpft.
Intestinales Immunsystem
Neben der Darmflora und der Epithelbarriere ist das intestinale Immunsystem die dritte wesentliche Komponente im Darm. Der Darm hat die meisten immunkompetenten Zellen des Körpers. Das intestinale Immunsystem (gut associated lymphoid tissue [GALT]) läuft entlang des gesamten Darmtrakts, umfasst die peyerschen Platten des Dünndarms, den Blinddarm und Follikel im Dickdarm (8). Dieses System enthält etwa zwei Drittel aller Lymphozyten, die sich vor allem unter dem Epithel in der Lamina pro-
Nr. 1 • 2008
17
Entzündung und Ernährung
pria und im Darmepithel befinden, und produziert die meisten Immunglobuline (vor allem sekretorisches IgA).
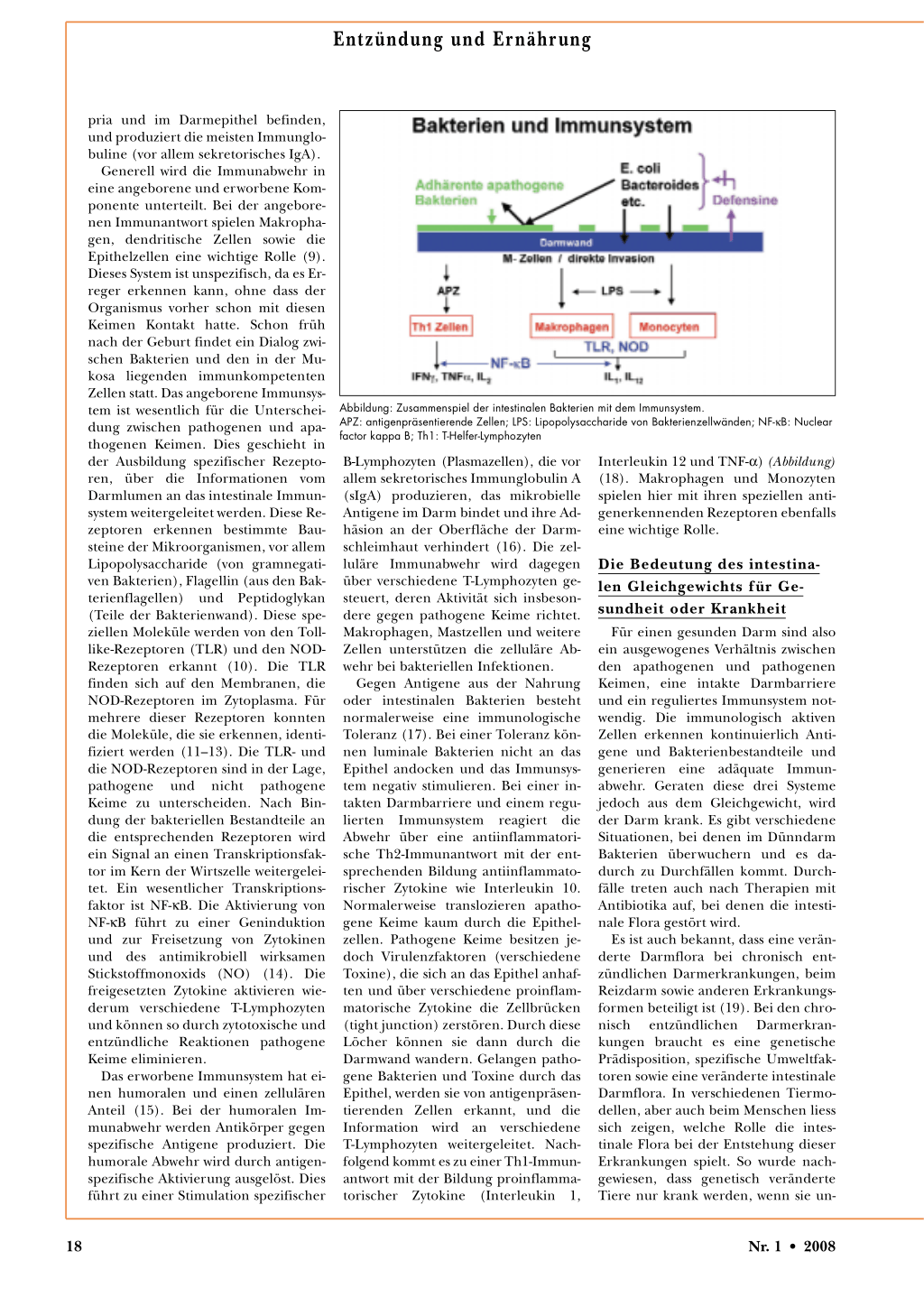
Generell wird die Immunabwehr in eine angeborene und erworbene Komponente unterteilt. Bei der angeborenen Immunantwort spielen Makrophagen, dendritische Zellen sowie die Epithelzellen eine wichtige Rolle (9). Dieses System ist unspezifisch, da es Erreger erkennen kann, ohne dass der Organismus vorher schon mit diesen Keimen Kontakt hatte. Schon früh nach der Geburt findet ein Dialog zwischen Bakterien und den in der Mukosa liegenden immunkompetenten Zellen statt. Das angeborene Immunsystem ist wesentlich für die Unterscheidung zwischen pathogenen und apathogenen Keimen. Dies geschieht in der Ausbildung spezifischer Rezeptoren, über die Informationen vom Darmlumen an das intestinale Immunsystem weitergeleitet werden. Diese Rezeptoren erkennen bestimmte Bausteine der Mikroorganismen, vor allem Lipopolysaccharide (von gramnegativen Bakterien), Flagellin (aus den Bakterienflagellen) und Peptidoglykan (Teile der Bakterienwand). Diese speziellen Moleküle werden von den Tolllike-Rezeptoren (TLR) und den NODRezeptoren erkannt (10). Die TLR finden sich auf den Membranen, die NOD-Rezeptoren im Zytoplasma. Für mehrere dieser Rezeptoren konnten die Moleküle, die sie erkennen, identifiziert werden (11–13). Die TLR- und die NOD-Rezeptoren sind in der Lage, pathogene und nicht pathogene Keime zu unterscheiden. Nach Bindung der bakteriellen Bestandteile an die entsprechenden Rezeptoren wird ein Signal an einen Transkriptionsfaktor im Kern der Wirtszelle weitergeleitet. Ein wesentlicher Transkriptionsfaktor ist NF-κB. Die Aktivierung von NF-κB führt zu einer Geninduktion und zur Freisetzung von Zytokinen und des antimikrobiell wirksamen Stickstoffmonoxids (NO) (14). Die freigesetzten Zytokine aktivieren wiederum verschiedene T-Lymphozyten und können so durch zytotoxische und entzündliche Reaktionen pathogene Keime eliminieren.
Das erworbene Immunsystem hat einen humoralen und einen zellulären Anteil (15). Bei der humoralen Immunabwehr werden Antikörper gegen spezifische Antigene produziert. Die humorale Abwehr wird durch antigenspezifische Aktivierung ausgelöst. Dies führt zu einer Stimulation spezifischer
Abbildung: Zusammenspiel der intestinalen Bakterien mit dem Immunsystem. APZ: antigenpräsentierende Zellen; LPS: Lipopolysaccharide von Bakterienzellwänden; NF-κB: Nuclear factor kappa B; Th1: T-Helfer-Lymphozyten
B-Lymphozyten (Plasmazellen), die vor allem sekretorisches Immunglobulin A (sIgA) produzieren, das mikrobielle Antigene im Darm bindet und ihre Adhäsion an der Oberfläche der Darmschleimhaut verhindert (16). Die zelluläre Immunabwehr wird dagegen über verschiedene T-Lymphozyten gesteuert, deren Aktivität sich insbesondere gegen pathogene Keime richtet. Makrophagen, Mastzellen und weitere Zellen unterstützen die zelluläre Abwehr bei bakteriellen Infektionen.
Gegen Antigene aus der Nahrung oder intestinalen Bakterien besteht normalerweise eine immunologische Toleranz (17). Bei einer Toleranz können luminale Bakterien nicht an das Epithel andocken und das Immunsystem negativ stimulieren. Bei einer intakten Darmbarriere und einem regulierten Immunsystem reagiert die Abwehr über eine antiinflammatorische Th2-Immunantwort mit der entsprechenden Bildung antiinflammatorischer Zytokine wie Interleukin 10. Normalerweise translozieren apathogene Keime kaum durch die Epithelzellen. Pathogene Keime besitzen jedoch Virulenzfaktoren (verschiedene Toxine), die sich an das Epithel anhaften und über verschiedene proinflammatorische Zytokine die Zellbrücken (tight junction) zerstören. Durch diese Löcher können sie dann durch die Darmwand wandern. Gelangen pathogene Bakterien und Toxine durch das Epithel, werden sie von antigenpräsentierenden Zellen erkannt, und die Information wird an verschiedene T-Lymphozyten weitergeleitet. Nachfolgend kommt es zu einer Th1-Immunantwort mit der Bildung proinflammatorischer Zytokine (Interleukin 1,
Interleukin 12 und TNF-α) (Abbildung) (18). Makrophagen und Monozyten spielen hier mit ihren speziellen antigenerkennenden Rezeptoren ebenfalls eine wichtige Rolle.
Die Bedeutung des intestina-
len Gleichgewichts für Ge-
sundheit oder Krankheit
Für einen gesunden Darm sind also ein ausgewogenes Verhältnis zwischen den apathogenen und pathogenen Keimen, eine intakte Darmbarriere und ein reguliertes Immunsystem notwendig. Die immunologisch aktiven Zellen erkennen kontinuierlich Antigene und Bakterienbestandteile und generieren eine adäquate Immunabwehr. Geraten diese drei Systeme jedoch aus dem Gleichgewicht, wird der Darm krank. Es gibt verschiedene Situationen, bei denen im Dünndarm Bakterien überwuchern und es dadurch zu Durchfällen kommt. Durchfälle treten auch nach Therapien mit Antibiotika auf, bei denen die intestinale Flora gestört wird.
Es ist auch bekannt, dass eine veränderte Darmflora bei chronisch entzündlichen Darmerkrankungen, beim Reizdarm sowie anderen Erkrankungsformen beteiligt ist (19). Bei den chronisch entzündlichen Darmerkrankungen braucht es eine genetische Prädisposition, spezifische Umweltfaktoren sowie eine veränderte intestinale Darmflora. In verschiedenen Tiermodellen, aber auch beim Menschen liess sich zeigen, welche Rolle die intestinale Flora bei der Entstehung dieser Erkrankungen spielt. So wurde nachgewiesen, dass genetisch veränderte Tiere nur krank werden, wenn sie un-
18 Nr. 1 • 2008
Entzündung und Ernährung
ter normalen Bedingungen mit bakteriellem Kontakt aufgezogen werden, bei steriler Aufzucht bleiben sie dagegen gesund (20–22). Liegt eine genetische Disposition vor, wird die Schutzwirkung der Darmbarriere aufgehoben, sodass pathogene Keime, über die Bindung an die spezifischen Rezeptoren (TLR, NOD), eine unkontrollierte Stimulation des Transkriptionsfaktors NF-κB mit vermehrter Bildung proinflammatorischer Zytokine (TNF-α, Interferon-γ, Interleukin 1, Interleukin 6 und Interleukin 12) auslösen können (23). Auch beim Reizdarm gibt es Hinweise, dass bei einem Teil der Patienten chronisch entzündliche Veränderungen im Darm vorliegen, an denen die intestinale Flora mit beteiligt ist (24–27). Wie Biopsien von Patienten mit Reizdarm gezeigt haben, unterscheidet sich die bakterielle Zusammensetzung der intestinalen Flora von jener gesunder Menschen (28).
Effekte von Prä- und Probio-
tika auf die Darmflora
Seit den Arbeiten von Ilja Metchnikoff, der 1908 den Nobelpreis erhalten hat, stand die Frage im Raum, ob sich das intestinale Ökosystem des Darms beeinflussen lässt, um gesundheitsfördernde, präventive oder therapeutische Effekte zu erzielen. Dies wurde in den letzten Jahren intensiv untersucht. Heute weiss man, dass sowohl spezifische Oligosaccharide (Präbiotika) als auch spezielle apathogene Bakterien (Probiotika) das intestinale System beeinflussen können.
Präbiotika Präbiotika haben die Fähigkeit, die
Darmflora positiv zu verändern. So konnte beispielsweise nach Gabe von Fructo-Oligosacchariden gezeigt werden, dass die Zahl der Bifidobakterien stark zunahm, während sich aerobe Keime, coliforme, grampositive Kokken und anaerobe Keime nur unwesentlich veränderten (29–31). Präbiotika werden bevorzugt durch Fermentationsprozesse von den ansässigen Bakterien zu kurzkettigen Fettsäuren gespalten. Durch die Fermentation kommt es zum Absinken des luminalen pH und dadurch auch zu einer Hemmung des Wachstums pathogener Keime. Ein Teil der kurzkettigen Fettsäuren (v.a. Butyrat) dient als Energiequelle für die Darmepithelzellen, aber auch als Nährsubstrat für die apathogenen Keime (v.a. Bifido-
bakterien und Lactobazillen) (32). Da sich diese Bakterien an die Schleimhaut des Darmepithels anhaften können, dienen sie auch zum Schutz vor der Invasion pathogener Keime in die Darmwand.
Probiotika Probotika sind spezifisch definierte
Mikroorganismen, für die eine Reihe von positiven Effekten im Darm nachgewiesen wurde. Probiotika regulieren die Darmbarriere und beeinflussen das intestinale Immunsystem. Sie sind in der Lage, proinflammatorische Zytokine zu hemmen und antiinflammatorische Zytokine zu stimulieren. Probiotika hemmen auch die Adhärenz von pathogenen Mikroorganismen an das Darmepithel sowie die Vermehrung pathogener gramnegativer, anaerober Keime durch die Sekretion von Bakteriozinen und blockieren die Translokation pathogener Keime durch das Darmepithel, gleichzeitig stimulieren sie das sIgA (19). Heute finden sich probiotische Bakterien wie Lactobazillen und Bifidobakterien in einer grossen Zahl von Milchprodukten. Darüber hinaus wurden aber auch weitere spezielle Bakterienstämme in klinischen Studien getestet. Die bestuntersuchten Bakterien für diese Zwecke sind E. coli Nissle 1917, Lactobacillus plantarum, L. reuteri, L. casei defensis, L. johnsonii, L. rhamnosus GG und verschiedene Bifidobakterien (33). Aufgrund der Tatsache, dass Präund Probiotika die Darmflora positiv beeinflussen, wurde ihnen ein genereller gesundheitsfördernder Effekt zugesprochen.
Prä- und Probiotika in Präven-
tion und Therapie von Krank-
heiten
In Europa werden Nahrungsmitteln vor allem Inulin und Fructo-Oligosaccharide beigefügt. Aufgrund der antiproliferativen Wirkungen der kurzkettigen Fettsäuren wird ein präventiver Effekt auf das Auftreten von Adenomen und Karzinomen im Dickdarm diskutiert. Diese Wirkung ist aber leider noch nicht eindeutig bewiesen, da keine guten Langzeitstudien beim Menschen einen solchen Effekt bis heute bestätigt haben (32). Ebenfalls gibt es nur wenig klinische Studien mit Präbiotika. Eine kleinere Studie zeigte mit Inulin einen positiven Effekt bei der akuten Pouchitis (Entzündung im Dünndarm nach einer totalen Dick-
darmentfernung) (34). Bei Kindern wurden nach Gabe von Fructo-Oligosacchariden gegenüber Plazebo weniger verstopfungsassoziierte Symptome gefunden. In der supplementierten Gruppe nahm auch die mittlere Stuhlfrequenz gegenüber Plazebo zu (35). Eine grössere Studie zeigte bei der Clostridium-difficile-assoziierten Diarrhö, dass die Rezidivraten nach Einsatz von Fructo-Oligosacchariden in der präbiotischen Gruppe geringer waren als mit Plazebo (36). Leider sind auch die propagierten gesundheitsfördernden Effekte für Probiotika bei der breiten Bevölkerung schlecht dokumentiert. Es gibt einige Hinweise, dass die regelmässige Einnahme von Probiotika bei älteren Menschen die natürliche Abwehr stärkt, was mit einer niedrigeren Infektionsrate einhergeht (37).
Für einige Probiotika wurde dagegen in klinischen Studien eine positive Wirkung bei verschiedenen gastrointestinalen Krankheiten gezeigt. Aufgrund dieser Studien können Probiotika heute bei Colitis ulcerosa durchaus zur Rezidivprophylaxe empfohlen werden (38, 39), bei der Pouchitis zur Prävention und Rezidivprophylaxe (40–42) sowie bei Durchfallerkrankungen zur Therapie und Prophylaxe (43). Eine gute Wirkung wurde vor allem in der Prophylaxe von antibiotikainduzierten Durchfällen gefunden (44–46). Bei Patienten mit Reizdarm kam es nach Probiotikagabe zu einer verbesserten Stuhlregulation und zu einer Abnahme der Blähungen (19, 28, 47). Bei einigen Erkrankungen wurden einzelne Probiotika oder Gemische verschiedener Probiotika untersucht. Für beide Verabreichungen konnten positive Effekte dokumentiert werden (19). Ob aber mehrere Probiotika nötig sind, ist zurzeit unklar.
Aufgrund der synergistischen Wirkungen der Prä- und Probiotika scheint es sinnvoll zu sein, beide Arten in Zukunft in Kombination als Symbiotika einzusetzen. Für Symbiotika wurde in einzelnen Gebieten wie beispielsweise bei der grossen Abdominalchirurgie sowie bei Lebertransplantationen ein positiver Effekt gezeigt, in dem weniger infektiöse Komplikationen auftraten (48, 49).
Fazit
Zusammenfassend sind die Erkenntnisse des Zusammenspiels der Prä- und Probiotika mit den intestinalen Mikro-
Nr. 1 • 2008
19
Entzündung und Ernährung
organismen der Darmbarriere und
dem intestinalen Immunsystem sehr
spannend. Wir befinden uns hier je-
doch erst am Anfang. Obwohl bereits
einige hochinteressante Aspekte be-
kannt sind, gibt es noch viele Unklar-
heiten. Da mit der Zufuhr von Prä- und
Probiotika das intestinale Milieu verän-
dert werden kann, sollten diese Pro-
dukte weiter zur Prävention und zur
Therapie von Erkrankungen in adä-
quaten Studien geprüft werden. Bei ei-
nigen Erkrankungen ist der Nutzen
gut belegt, für andere aber bestehen
lediglich Hinweise mit teilweise kon-
troversen Daten. Unklar ist beispiels-
weise, welche Produkte sich am besten
eignen, welche Mengen und wie lange
sie verabreicht werden müssen. Diese
Fragen müssen noch gelöst werden,
damit der Einsatz von Prä- und Probio-
tika empfohlen werden kann.
I
Korrespondenzadresse: Prof. Dr. med. Rémy Meier Leitender Arzt Gastroenterologie, Hepatologie und Ernährung Medizinische Universitätsklinik Kantonsspital Liestal 4410 Liestal Tel. 061-925 21 87 Fax 061-925 28 04 E-Mail: remy.meier@ksli.ch
Referenzen: 1. Bengmark S. Colonic food: pre- and probiotics. Am J Gastroenterol 2000; 95: S5–S7. 2. Sartor RB. Therapeutic manipulation of the enteric microflora in inflammatory bowel diseases: antibiotics, probiotics, and prebiotics. Gastroenterology 2004; 126: 1620–1633. 3. Lievin V, Pfeiffer I, Hudault S, et al. Bifidobacterium strains from resident infant human gastrointestinal microflora exert antimicrobial acitvity. Gut 2000;47: 646–652. 4. Inan MS, Rasoulpour RJ, Yin L, et al. The luminal short-chain fatty acid butyrate modulates NFκB activity in a human colonic epithelial cell line. Gastroenterology 2000; 118: 724–734. 5. Wachtershauser A, Loitsch SM, Stein J. PPARgamma is selectively upregulated in Caco-2 cells by butyrate. Biochem Biophys Res Commun 2000; 272: 380–385. 6. Brandtzaeg P, Pabst R. Let’s go mucosal: communication on slippery ground. Trends Immunol 2004; 25: 570–577. 7. Pabst R. Mukosa-assoziiertes lymphatisches Gewebe des Darms. In: Drenckhahn D (Hrsg): Benninghoff-Drenckhahn Anatomie. Band II, 16. Auflage. München: Elsevier Urban & Fischer, 2004: 165–167. 8. Brandtzaeg P, Halstensen TS, Kett K, et al. Immunobiology and immunopathology of human gut mucosa: humoral immunity and intraepithelial lymphocytes. Gastroenterology 1989; 97: 1562–1584. 9. Rimoldi M, Chieppa M, Salucci V, et al. Intestinal immune homeostasis is regulated by the crosstalk between epithelial cells and dendritic cells. Nature Immunol 2005; 6: 507–514. 10. Eckmann L. Innate immunity and mucosal bacterial interactions in the intestine. Curr Opin Gastroenterol 2004; 20: 82–88.
11. Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, et al. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 2004; 118: 229–241. 12. Abreu MT, Fukata M, Arditi M. TLR signalling in the gut in health and disease. J Immunol 2005; 174: 4453–4460. 13. Chamaillard M, Inohara N, Nunez G. Batteling enteroinvasive bacteria: Nod1 comes to the rescue. Trends Microbiol 2004; 12: 529–532. 14. Neish AS, Gewirtz AZ, Teng H, et al. Prokaryotic regulation of epithelial responses by inhibition of IκB-α-ubiquitination. Science 2000; 289: 1560–1563. 15. Delves PJ, Roitt IM. The Immune System. N Engl J Med 2000; 343: 37–49. 16. Kagnoff MF. Immunology of the intestinal tract. Gastroenterology 1993; 105: 1275– 1280. 17. Mowat AMCI. Anatomical basis of tolerance and immunity to intestinal antigens. Nature reviews 2003; 3: 331–341. 18. Sartor RB. Induction of mucosal immune responses by bacteria and bacterial components. Current Opinion in Gastroenterology 2001; 17: 555–561. 19. Meier R. Probiotika. Die Bedeutung der intestinalen Flora für die Genese und Therapie gastrointestinaler Erkrankungen. Schweiz Med Forum 2007; 7: 184–191. 20. Sadlack B, Merz H, Schorle H, et al. Ulcerative colitis-like disease in mice with a disrupted interleukin-2 gene. Cell Microbiol 1993; 75: 253–261. 21. Kuhn R, Löhler J, Rennick D, et al. Interleukin10-deficient mice develop chronic enterocolitis. Cell 1993; 75: 263–274. 22. Madsen KL, Doyle JS, Jewell LD, et al. Lactobacillus species prevents colitis in interleukin 10 gene-deficient mice. Gastroenterology 1999; 116: 1107–1114. 23. Mahida YR, Rolfe VE. Host-bacterial interactions in inflammatory bowel disease. Clin Sci (Lond) 2004; 107 (4): 331–341. 24. Spiller RC. Neuropathology of IBS? Gastroenterology 2002; 123: 2144–2147. 25. Balsari A, Ceccarelli A, Dubini F, et al. The fecal microbial population in the irritable bowel syndrome. Microbiologica 1982; 5: 185–194. 26. Nobaek S, Johansson ML, Molin G, et al. Alteration of intestinal microflora is associated with reduction in abdominal bloating and pain in patients with irritable bowel syndrome. Am J Gastroenterol 2000; 95: 1231–1238. 27. O’Mahony L, McCarthy J, Kelly P, et al. Lactobacillus and bifidobacterium in irritable bowel syndrome: symptom responses and relationship to cytokine profiles. Gastroenterology 2005; 128 (3): 541–551. 28. Swidsinski A, Khilkin M, Ortner M, et al. Alteration of bacterial concentration in colonic biopsies from patients with irritable bowel syndrome (IBS). Gastroenterology 1999; 116: A1. 29. Wang X, Gibson GR. Effects of the in vitro fermentation of oligofructose and inulin by bacteria growing in the human large intestine. J Appl Bacteriol 1993; 75: 373–80. 30. Gibson GR, Wang X. Regulatory effects of bifidobacteria on the growth of other colonic bacteria. J Appl Bacteriol 1994; 77: 412–420. 31. Gibson GR, Beatty ER, Wang X, et al. Selective stimulation of bifidobacteria in the human colon by oligofructose and inulin. Gastroenterology 1995; 108: 975–982. 32. Scheppach W, Weiler F. The butyrate story: old wine in new bottles? Curr Opin Clin Nutr Metab Care 2004; 7: 563–567. 33. Meier R. Lochs H. Prä- und Probiotika. Therapeutische Umschau 2007; 64: 161–169.
34. Welters CF, Heineman E, Thunnissen FB, et al. Effect of dietary inulin supplementation on inflammation of pouch mucosa in patients with an ileal pouch-anal anastomosis. Dis Colon Rectum 2002; 45: 621–627. 35. Moore N, Chao C, Yang LP, et al. Effects of fructo-oligosaccharide-supplemented infant cereal: a double blind, randomized trial. Br J Nutr 2003; 90: 581–587. 36. Lewis S, Burmeister S, Brazier J. Effect of the prebiotic oligofructose on relapse of Clostridium difficile-associated diarrhea: a randomized, controlled study. Clin Gastroenterol Hepatol 2005; 3: 442–448. 37. Bunout D, Barrera G, Hirsch S, et al. Effects of a nutritional supplement on the immune response and cytokine production in free-living Chilean elderly. J Parenter Enteral Nutr 2004; 28: 348–354. 38. Kruis W, Fric P, Pokrotnieks J, et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut 2004; 53: 1617–23. 39. Zocco MA, Zileri dal Verme L, Cremonini F, et al. Efficacy of Lactobacillus GG in maintaining remission of ulcerative colits. Aliment Pharmacol Ther 2006; 23: 1567–1574. 40. Gionchetti P, Rizzello F, Venturi A, et al. Oral bacteriotherapy as maintenance treatment in patients with chronic pouchitis: a double-blind, placebo-controlled trial. Gastroenterology 2000; 119: 305–309. 41. Gionchetti P, Rizzello F, Helwig U, et al. Prophylaxis of pouchitis onset with probiotic therapy: a double-blind, placebo-controlled trial. Gastroenterology 2003; 124: 1202–1209. 42. Mimura T, Rizzello F, Helwig U, et al. Once daily high dose probiotic therapy (VSL#3) for maintaining remission in recurrent or refractory pouchitis. Gut 2004; 53: 108–114. 43. Szajewska H, Mrukowicz JZ. Probiotics in the treatment and prevention of acute infectious diarrhea in infants and children: a systematic review of published randomized, double-blind, placebo-controlled trials. J Pediatr Gastroenterol Nutr 2001; 33 (Suppl 2): S17–S25. 44. D’Souza AL, Rajkumar C, Cooke J, et al. Probiotics in prevention of antibiotic associated diarrhoea: meta-analysis. BMJ 2002; 324: 1361. 45. Cremonini F, Di Caro S, Nista EC, et al. Meta-analysis: the effect of probiotic administration on antibiotic-associated diarrhoea. Aliment Pharmacol Ther 2002; 16: 1461–1467. 46. Allen SJ, Okoko B, Martinez E, et al. Probiotics for treating infectious diarrhoea. Cochrane Database Syst Rev 2004; 2: CD003048. 47. Bazzocchi G, Almerigi PF, Gionchetti P, Campieri M. Changes in symptoms, distensionstimulated colonic motility and in fecal microbiological features after oral bacteriotherapy in patients with diarrhea-predominant IBS (D-IBS) with functional diarrhea (FD). Am J Gastroenterol 2002; 97: A847. 48. Rayes N, Hansen S, Seehofer D, et al. Early enteral supply of fiber and Lactobacilli versus conventional nutrition: a controlled trial in patients with major abdominal surgery. Nutrition 2002; 18: 609–615. 49. Rayes N, Seehofer D, Theruvath T, et al. Effect of enteral nutrition and synbiotics on bacterial infection rates after pylorus-preserving pancreatoduodenectomy. A randomized, double-blind trial. Ann Surg 2007; 246: 36–41.
20 Nr. 1 • 2008